Balik Clinic Oxidative Stress Chiropractic ug Functional Medicine Team. Ang stress sa oxidative gihubit isip usa ka kasamok sa balanse tali sa paghimo sa reaktibo nga oxygen (free radicals) ug mga depensa sa antioxidant. Sa laing pagkasulti, kini usa ka dili balanse tali sa paghimo sa mga libre nga radikal ug ang abilidad sa lawas sa pagsumpo o pag-detoxify sa makadaot nga mga epekto pinaagi sa pag-neutralize sa mga antioxidant. Ang stress sa oxidative nagdala sa daghang mga kondisyon sa pathophysiological sa lawas. Kini naglakip sa neurodegenerative nga mga sakit, ie, Parkinson's disease, Alzheimer's disease, gene mutation, cancers, chronic fatigue syndrome, fragile X syndrome, heart and blood vessel disorders, atherosclerosis, heart failure, heart attack, ug inflammatory disease. Ang oksihenasyon mahitabo ubos sa daghang mga kahimtang:
ang mga selyula naggamit sa glucose sa paghimo og enerhiya
ang immune system nakig-away sa bakterya ug nagmugna og panghubag
ang mga lawas nag-detoxize sa mga pollutant, pestisidyo, ug aso sa sigarilyo
Adunay minilyon nga mga proseso nga nahitabo sa atong mga lawas sa bisan unsang oras nga mahimong moresulta sa oksihenasyon. Ania ang pipila ka mga sintomas:
kakapoy
Pagkawala sa memorya ug o fog sa utok
Kaunuran ug o lutahan kasakit
Mga kunot kauban ang ubanon nga buhok
Pagkunhod sa panan-aw
Sakit sa ulo ug pagkasensitibo sa kasaba
Pagdugay sa mga impeksyon
Ang pagpili sa mga organikong pagkaon ug paglikay sa mga hilo sa imong palibot dakog kalainan. Kini, uban sa pagkunhod sa tensiyon, mahimong mapuslanon sa pagkunhod sa oksihenasyon.
Ang mga oxidant sa kasagaran gihimo sa usa ka kontrolado nga paagi aron makontrol ang hinungdanon nga mga proseso sa lawas sa tawo, lakip ang cell division, panghubag, immune function, autophagy, ug pagtubag sa stress. Bisan pa, ang dili makontrol nga produksiyon niini nga mga oxidant mahimong makatampo sa oxidative stress, nga mahimong makaapekto sa cellular function, nga modala ngadto sa kalamboan sa toxicity, laygay nga sakit ug kanser. Ang mga mekanismo sa pagpanalipod sa antioxidant sa lawas sa tawo gi-regulate sa usa ka serye sa hinungdanon nga mga agianan nga nagkontrol sa tubag sa cell sa mga oxidant. Ang nukleyar nga hinungdan nga erythroid 2-related nga butang, o nailhan nga Nrf2, usa ka nag-uswag nga regulator sa cellular resistance sa mga oxidant. Ang katuyoan sa artikulo sa ubos mao ang paghisgot ug pagpakita sa mitumaw nga papel sa Nrf2 sa mitochondrial function.
abstract
Ang transcription factor NF-E2 p45-related factor 2 (Nrf2; gene name NFE2L2) nagtugot sa pagpahiangay ug pagkaluwas ubos sa mga kondisyon sa stress pinaagi sa pag-regulate sa gene nga ekspresyon sa lain-laing mga network sa cytoprotective proteins, lakip na ang antioxidant, anti-inflammatory, ug detoxification enzymes usab isip mga protina nga nagtabang sa pag-ayo o pagtangtang sa nadaot nga mga macromolecule. Ang Nrf2 adunay importante nga papel sa pagmentinar sa cellular redox homeostasis pinaagi sa pag-regulate sa biosynthesis, paggamit, ug pagbag-o sa glutathione, thioredoxin, ug NADPH ug pinaagi sa pagkontrolar sa produksyon sa reactive oxygen species pinaagi sa mitochondria ug NADPH oxidase. Ubos sa mga kondisyon sa homeostatic, ang Nrf2 makaapekto sa potensyal sa mitochondrial membrane, fatty acid oxidation, pagkabaton sa substrates (NADH ug FADH2 / succinate) alang sa respiration, ug ATP synthesis. Ubos sa mga kondisyon sa stress o growth factor stimulation, ang pagpaaktibo sa Nrf2 mosumpo sa dugang nga reactive oxygen species nga produksyon sa mitochondria pinaagi sa transcriptional upregulation sa uncoupling protein 3 ug nag-impluwensya sa mitochondrial biogenesis pinaagi sa pagpadayon sa lebel sa nuclear respiratory factor 1 ug peroxisome proliferator-activated receptor? coactivator 1?, ingon man pinaagi sa pagpasiugda sa purine nucleotide biosynthesis. Ang mga activator sa Pharmacological Nrf2, sama sa natural nga isothiocyanate sulforaphane, nagpugong sa oxidant-mediated nga pag-abli sa mitochondrial permeability transition pore ug mitochondrial swelling. Katingad-an, ang usa ka sintetikong 1,4-diphenyl-1,2,3-triazole compound, nga orihinal nga gidisenyo isip Nrf2 activator, nakit-an nga nagpasiugda sa mitophagy, sa ingon nakatampo sa kinatibuk-ang mitochondrial homeostasis. Busa, ang Nrf2 usa ka prominenteng magdudula sa pagsuporta sa structural ug functional nga integridad sa mitochondria, ug kini nga papel ilabinang mahinungdanon ubos sa mga kondisyon sa tensiyon.
Ang Nrf2 adunay hinungdanon nga papel sa pagpadayon sa cellular redox homeostasis.
Ang Nrf2 makaapekto sa potensyal sa mitochondrial membrane ug ATP synthesis.
Ang Nrf2 nag-impluwensya sa mitochondrial fatty acid oxidation.
Gisuportahan sa Nrf2 ang structural ug functional nga integridad sa mitochondria.
Ang Nrf2 activators adunay mapuslanon nga mga epekto kung ang mitochondrial function makompromiso.
Pasiuna
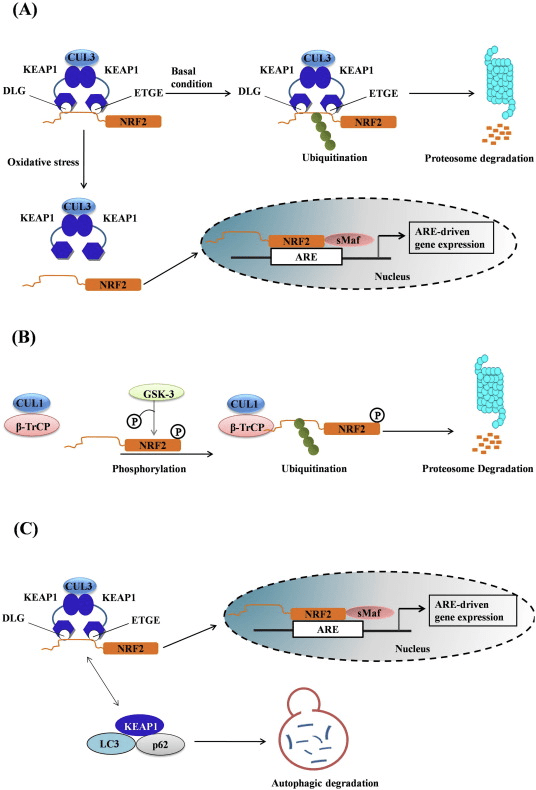
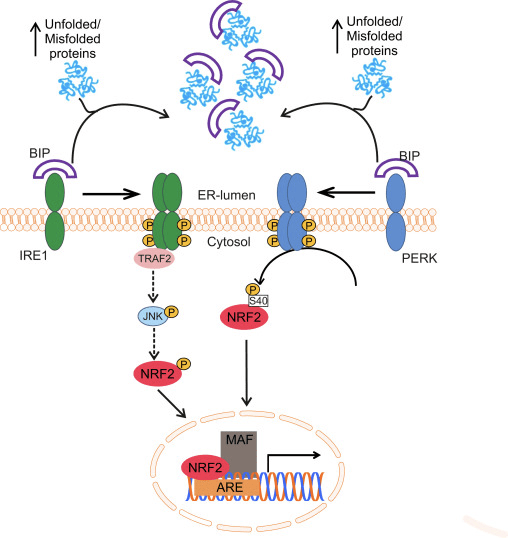
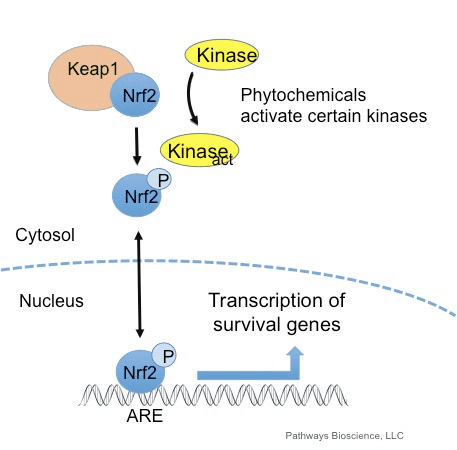
Ang transcription factor NF-E2 p45-related factor 2 (Nrf2; gene name NFE2L2) nag-regulate sa pagpahayag sa mga network sa mga gene nga nag-encode sa mga protina nga adunay lainlain nga cytoprotective nga mga kalihokan. Ang Nrf2 mismo kontrolado una sa lebel sa kalig-on sa protina. Ubos sa basal nga mga kondisyon, ang Nrf2 usa ka mubo nga kinabuhi nga protina nga gipailalom sa padayon nga ubiquitination ug proteasomal degradation. Adunay tulo ka nahibal-an nga ubiquitin ligase system nga nakatampo sa pagkadaot sa Nrf2. Sa kasaysayan, ang unang negatibo nga regulator sa Nrf2 nga nadiskobrehan mao ang Kelch-like ECH-associated protein 1 (Keap1) [1], usa ka substrate adapter protein alang sa Cullin 3 (Cul3) / Rbx1 ubiquitin ligase [2], [3], [ 4]. Ang Keap1 naggamit sa usa ka episyente kaayo nga cyclic nga mekanismo sa pag-target sa Nrf2 alang sa ubiquitination ug proteasomal degradation, diin ang Keap1 padayon nga gibag-o, nga nagtugot sa cycle nga magpadayon (Fig. 1A) [5]. Ang Nrf2 gipailalom usab sa degradasyon nga gipataliwad-an sa glycogen synthase kinase (GSK) 3/?-TrCP-dependent Cul1-based ubiquitin ligase [6], [7]. Labing bag-o lang, gitaho nga, sa panahon sa mga kondisyon sa endoplasmic reticulum stress, ang Nrf2 kay ubiquitinated ug degraded sa usa ka proseso nga gipataliwala sa E3 ubiquitin ligase Hrd1 [8].
Figure 1 Ang cyclic sequential binding ug regeneration model alang sa Keap1-mediated degradation sa Nrf2. (A) Ang Nrf2 nagbugkos nga sunud-sunod sa usa ka libre nga Keap1 dimer: una pinaagi sa iyang high-affinity ETGE (pula nga sticks) binding domain ug dayon pinaagi sa iyang low-affinity DLG (black sticks) binding domain. Niini nga pagpahiuyon sa komplikadong protina, ang Nrf2 nakaagi sa ubiquitination ug gipunting alang sa pagkadaot sa proteasomal. Ang Free Keap1 nabag-o ug makahimo sa pagbugkos sa bag-ong gihubad nga Nrf2, ug ang siklo nagsugod pag-usab.(B) Ang mga inducers (puti nga diamante) mo-react sa sensor cysteinees sa Keap1 (asul nga mga stick), nga mitultol sa usa ka conformational nga pagbag-o ug ningdaot nga substrate adapter nga kalihokan. Ang libre nga Keap1 wala gibag-o, ug ang bag-ong gi-synthesize nga Nrf2 natipon ug gibalhin sa nucleus.
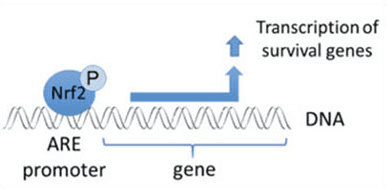
Dugang sa pagserbisyo isip ubiquitin ligase substrate adapter protein, ang Keap1 mao usab ang sensor alang sa usa ka halapad nga han-ay sa gagmay nga molekula nga mga activator sa Nrf2 (gitawag nga mga inducers) [9]. Gibabagan sa mga inducers ang siklo sa Keap1-mediated degradation sa Nrf2 pinaagi sa kemikal nga pagbag-o sa piho nga mga residu sa cysteine sulod sa Keap1 [10], [11] o pinaagi sa direktang pagsamok sa Keap1: Nrf2 binding interface [12], [13]. Tungod niini, ang Nrf2 dili degraded, ug ang transcription factor natipon ug gibalhin ngadto sa nucleus (Fig. 1B), diin kini nagporma og heterodimer nga adunay gamay nga protina sa Maf; nagbugkos sa antioxidant-response nga mga elemento, ang upstream regulatory regions sa iyang target nga mga gene; ug gisugdan ang transkripsyon [14], [15], [16]. Ang baterya sa mga target sa Nrf2 naglangkob sa mga protina nga adunay lainlaing mga function sa cytoprotective, lakip ang mga enzyme sa xenobiotic metabolism, mga protina nga adunay antioxidant ug anti-inflammatory function, ug proteasomal subunits, ingon man mga protina nga nag-regulate sa cellular redox homeostasis ug nag-apil sa intermediary metabolism.
Nrf2: usa ka Master Regulator sa Cellular Redox Homeostasis
Ang function sa Nrf2 ingon usa ka master regulator sa cellular redox homeostasis kaylap nga giila. Ang ekspresyon sa gene sa catalytic ug sa regulatory subunits sa ?-glutamyl cysteine ligase, ang enzyme nga nag-catalyze sa rate-limiting nga lakang sa biosynthesis sa pagkunhod sa glutathione (GSH), direktang gi-regulate sa Nrf2 [17]. Ang xCT subunit sa sistema xc-, nga nag-import sa cystine ngadto sa mga selula, usa usab ka direkta nga transcriptional nga target sa Nrf2 [18]. Sa selula, ang cystine moagi sa pagkakabig ngadto sa cysteine, usa ka pasiuna sa biosynthesis sa GSH. Dugang pa sa papel niini sa GSH biosynthesis, ang Nrf2 naghatag og paagi alang sa pagmentinar sa glutathione sa pagkunhod niini pinaagi sa coordinated transcriptional regulation sa glutathione reductase 1 [19], [20], nga nagpamenos sa oxidized glutathione ngadto sa GSH gamit ang pagkunhod sa mga katumbas gikan sa NADPH . Ang gikinahanglan nga NADPH gihatag sa upat ka punoan nga NADPH-generating enzymes, malic enzyme 1 (ME1), isocitrate dehydrogenase 1 (IDH1), glucose-6-phosphate dehydrogenase (G6PD), ug 6-phosphogluconate dehydrogenase (PGD), nga tanan transcriptionally regulated sa bahin sa Nrf2 (Fig. 2) [21], [22], [23], [24]. Katingad-an, ang Nrf2 usab nag-regulate sa inducible gene expression sa cytosolic, microsomal, ug mitochondrial nga mga porma sa aldehyde dehydrogenase [25], nga naggamit sa NAD (P) + isip usa ka cofactor, nga nagpatungha sa NAD (P) H. Sa tinuud, ang lebel sa NADPH ug ang NADPH / NADP + ratio mas ubos sa mga embryonic fibroblast nga nahimulag gikan sa Nrf2-knockout (Nrf2-KO) nga mga ilaga kumpara sa mga selula gikan sa ilang wild-type (WT) nga mga katugbang, ug ang lebel sa NADPH mikunhod sa Nrf2 knockdown sa mga linya sa selula sa kanser nga adunay aktibo nga Nrf2 [26]. Sama sa gipaabot, ang lebel sa GSH mas ubos sa mga selula diin ang Nrf2 nabalda; Sa kasukwahi, ang pagpaaktibo sa Nrf2 pinaagi sa genetic o pharmacological nga paagi nagdala ngadto sa GSH upregulation [27], [28], [29]. Importante, ang Nrf2 usab nag-regulate sa gene nga ekspresyon sa thioredoxin [30], [31], [32], thioredoxin reductase 1 [28], [29], [32], [33], ug sulfiredoxin [34], nga hinungdanon. alang sa pagkunhod sa oxidized protein thiols.
Figure 2 Ang papel sa Nrf2 sa metabolismo sa paspas nga pagdaghan sa mga selula. Ang Nrf2 usa ka positibo nga regulator sa mga gene nga nag-encode sa mga enzyme sa duha nga oxidative arm [ie, glucose-6-phosphate dehydrogenase (G6PD) ug 6-phosphogluconate dehydrogenase (PGD)] ug ang nonoxidative arm [ie, transaldolase 1 (TALDO1) ug transketolase ( TKT)] sa pentose phosphate nga agianan. G6PD ug PGD makamugna NADPH. Ang Nrf2 usab nag-regulate sa gene nga ekspresyon sa laing duha ka NADPH-generating enzymes, malic enzyme 1 (ME1) ug isocitrate dehydrogenase 1 (IDH1). Ang ekspresyon sa gene sa phosphoribosyl pyrophosphate amidotransferase (PPAT), nga nag-catalyze sa pagsulod sa de novo purine biosynthetic pathway, positibo usab nga gi-regulate sa Nrf2, sama sa ekspresyon sa methylenetetrahydrofolate dehydrogenase 2 (MTHFD2), usa ka mitochondrial enzyme nga adunay kritikal nga papel sa paghatag og usa ka-carbon nga mga yunit alang sa de novo purine biosynthesis. Ang Pyruvate kinase (PK) negatibo nga gikontrol sa Nrf2 ug gilauman nga mopabor sa pagtukod sa glycolytic intermediates ug, uban sa G6PD, metabolite channeling pinaagi sa pentose phosphate nga agianan ug ang synthesis sa nucleic acid, amino acids, ug phospholipids. Ang Nrf2 negatibo nga nagkontrol sa gene nga ekspresyon sa ATP-citrate lyase (CL), nga mahimong makadugang sa pagkaanaa sa citrate alang sa paggamit sa mitochondrial o (pinaagi sa isocitrate) alang sa IDH1. Ang pula ug asul nagpaila sa positibo ug negatibo nga regulasyon, matag usa. Ang mitochondrion gipakita sa gray. Metabolite abbreviation: G-6-P, glucose 6-phosphate; F-6-P, fructose 6-phosphate; F-1,6-BP, fructose 1,6-bisphosphate; GA-3-P, glyceraldehyde 3-phosphate; 3-PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; 6-P-Gl, 6-phosphogluconolactone; 6-PG, 6-phosphogluconate; R-5-P, ribulose 5-phosphate; PRPP, 5-phosphoribosyl-?-1-pyrophosphate; THF, tetrahydrofolate; IMP, inosine monophosphate; AMP, adenosine monophosphate; GMP, guanosine monophosphate.
Tungod sa mahinungdanong papel sa Nrf2 isip master regulator sa cellular redox homeostasis, dili ikatingala nga, kon itandi sa WT cells, ang lebel sa reactive oxygen species (ROS) mas taas sa mga selula diin ang Nrf2 nabalda (Nrf2-KO) [35]. Kini nga kalainan labi ka makapakurat sa hagit sa mga ahente nga hinungdan sa stress sa oxidative. Dugang pa, ang mga selula nga kulang sa Nrf2 mas sensitibo sa toxicity sa mga oxidant sa nagkalain-laing matang ug dili mapanalipdan sa Nrf2 inducers, nga, ubos sa sama nga mga kondisyon, naghatag og episyente ug dugay nga panalipod sa WT cells [29], [36] , [37]. Gawas pa sa kinatibuk-ang cellular redox homeostasis, ang Nrf2 kritikal usab alang sa pagpadayon sa mitochondrial redox homeostasis. Sa ingon, kung itandi sa WT, ang kinatibuk-ang mitochondrial NADH pool labi nga nadugangan sa Keap1-KO ug mahinuklugong mikunhod sa Nrf2-KO nga mga selula [35].
Gamit ang live cell imaging, bag-ohay lang nga gimonitor namon ang mga rate sa produksiyon sa ROS sa panguna nga glioneuronal cocultures ug mga hiwa sa tisyu sa utok nga nahimulag gikan sa WT, Nrf2-KO, o Keap1-knockdown (Keap1-KD) nga mga ilaga [38]. Sama sa gipaabut, ang rate sa produksiyon sa ROS mas paspas sa mga selula ug tisyu sa Nrf2-KO kumpara sa ilang mga katugbang sa WT. Bisan pa, gihimo namon ang wala damha nga obserbasyon nga, kung itandi sa WT, ang mga selula sa Keap1-KD adunay mas taas nga rate sa produksiyon sa ROS, bisan kung ang kadako sa kalainan tali sa WT ug Keap1-KD genotypes mas gamay kaysa sa taliwala sa WT ug Nrf2-KO . Gisusi dayon namo ang lebel sa mRNA sa NOX2 ug NOX4, ang mga catalytic subunits sa duha ka NADPH oxidase (NOX) isoforms nga nalambigit sa patolohiya sa utok, ug nakit-an nga ang NOX2 mahinuklugong nadugangan ubos sa kondisyon sa Nrf2 deficiency, samtang ang NOX4 gi-upregulated sa dihang Nrf2 constitutively activate, bisan pa sa usa ka gamay nga gidak-on. Sa quantitatively, ang kadako sa upregulation sa mga selula ug mga tisyu gikan sa mutant nga mga ilaga susama sa katugbang nga pagtaas sa produksyon sa ROS [38]. Makapainteres, dili lamang ang Nrf2 ang nag-regulate sa NADPH oxidase, apan ang ROS nga gihimo sa NADPH oxidase maka-activate sa Nrf2, ingon sa gipakita sa pulmonary epithelial cells ug cardiomyocytes [39], [40]. Dugang pa, ang usa ka bag-o nga pagtuon nagpakita nga ang NADPH oxidase-dependent activation sa Nrf2 naglangkob sa usa ka importante nga endogenous nga mekanismo alang sa panalipod batok sa mitochondrial nga kadaot ug cell death sa kasingkasing atol sa chronic pressure overload [41].
Dugang pa sa catalytic nga kalihokan sa NADPH oxidase, mitochondrial respiration mao ang lain nga mayor nga intracellular tinubdan sa ROS.Pinaagi sa paggamit sa mitochondria-specific probe MitoSOX, atong gisusi ang kontribusyon sa ROS sa mitochondrial gigikanan ngadto sa kinatibuk-ang ROS produksyon sa nag-unang glioneuronal cocultures nahimulag gikan sa WT, Nrf2-KO, o Keap1-KD nga mga ilaga [38]. Sama sa gipaabut, ang mga selula sa Nrf2-KO adunay mas taas nga rate sa produksiyon sa mitochondrial ROS kaysa WT. Sa pag-uyon sa mga nahibal-an alang sa kinatibuk-ang produksiyon sa ROS, ang mga rate sa produksiyon sa mitochondrial ROS sa Keap1-KD mas taas usab kung itandi sa mga selula sa WT. Importante, ang pag-ali sa complex I nga adunay rotenone hinungdan sa usa ka talagsaong pagtaas sa mitochondrial ROS nga produksyon sa WT ug Keap1-KD nga mga selula, apan walay epekto sa Nrf2-KO nga mga selula. Sukwahi sa gipaabot nga pagtaas sa produksyon sa mitochondrial ROS sa mga selula sa WT human sa pagdugang sa pyruvate (aron mapalambo ang pagkaanaa sa NADH, pagdugang sa potensyal sa mitochondrial membrane, ug pag-normalize sa respiration), ang produksyon sa ROS mikunhod sa Nrf2-KO nga mga selula. Mag-uban, kini nga mga nahibal-an kusganong nagsugyot nga, kung wala ang Nrf2: (i) ang kalihokan sa complex I nadaot, (ii) ang ningdaot nga kalihokan sa complex I tungod sa limitasyon sa mga substrate, ug (iii) ang ningdaot nga kalihokan sa komplikado. I mao ang usa sa mga nag-unang rason alang sa dugang nga mitochondrial ROS produksyon, lagmit tungod sa balihon electron dagan gikan sa complex II.
Ang Nrf2 Nakaapekto sa Potensyal ug Respirasyon sa Mitochondrial Membrane
Ang potensyal sa mitochondrial membrane (??m) usa ka unibersal nga timailhan sa kahimsog sa mitochondrial ug ang metaboliko nga kahimtang sa selula. Sa usa ka himsog nga selula, ??m gimentinar sa mitochondrial respiratory chain. Makapainteres, ang usa ka lig-on nga isotopic nga pag-label nga adunay mga amino acid sa pagtuon nga nakabase sa kultura nga proteomics sa estrogen receptor-negative nontumorigenic human breast epithelial MCF10A cell line nagpakita nga ang mitochondrial electron transport chain component NDUFA4 gi-upregulated sa pharmacological activation (sa sulforaphane) sa Nrf2, samtang ang genetic upregulation sa Nrf2 (sa Keap1 knockdown) mosangpot sa downregulation sa cytochrome c oxidase subunits COX2 ug COX4I1 [42]. Usa ka pagtuon sa proteome sa atay gamit ang two-dimensional gel electrophoresis ug matrix-assisted laser desorption/ionization mass spectrometry nakakaplag nga ang Nrf2 nag-regulate sa ekspresyon sa ATP synthase subunit? [43]. Dugang pa, ang mitochondrial protein DJ-1, nga adunay papel sa pagmentinar sa kalihokan sa complex I [44], gitaho nga nagpalig-on sa Nrf2 [45], [46], bisan kung ang neuroprotective nga epekto sa pharmacological o genetic activation sa Nrf2 independente sa DJ-1 [47]. Bisan pa, ang mga sangputanan niini nga mga obserbasyon alang sa mitochondrial function wala pa masusi.
Sa pag-uyon sa ningdaot nga kalihokan sa complex I ubos sa mga kondisyon sa Nrf2 deficiency, ang basal m mas ubos sa Nrf2-KO mouse embryonic fibroblasts (MEFs) ug kultura nga primary glioneuronal cells kon itandi sa ilang WT counterparts (Fig. 3, inset) [35]. Sa kasukwahi, ang basal m mas taas kung ang Nrf2 kay genetically constitutively upregulated (pinaagi sa knockdown o knockout sa Keap1). Kini nga mga kalainan sa ??m taliwala sa mga genotype nagpakita nga ang pagginhawa apektado sa kalihokan sa Nrf2. Sa pagkatinuod, ang pagtimbang-timbang sa konsumo sa oxygen sa basal nga estado nagpadayag nga, kon itandi sa WT, ang konsumo sa oxygen mas ubos sa Nrf2-KO ug Keap1-KO MEFs, sa ~ 50 ug ~ 35%, matag usa.
Figure 3 Gisugyot nga mekanismo alang sa nakompromiso nga mitochondrial function ubos sa kondisyon sa Nrf2 deficiency. (1) Ang pagkunhod sa lebel sa ME1, IDH1, G6PD, ug PGD moresulta sa ubos nga lebel sa NADPH. (2) Ang lebel sa GSH ubos usab. (3) Ang ubos nga kalihokan sa ME1 mahimong makapakunhod sa pool sa pyruvate nga mosulod sa mitochondria. (4) Ang henerasyon sa NADH mas hinay, nga nagdala ngadto sa ningdaot nga kalihokan sa complex I ug nagdugang sa produksyon sa mitochondrial ROS. (5) Ang pagkunhod sa FAD ngadto sa FADH2 sa mitochondrial proteins usab mikunhod, pagpaubos sa electron flow gikan sa FADH2 ngadto sa UbQ ug ngadto sa complex III. (6) Ang hinay nga pagporma sa UbQH2 mahimong magpaubos sa kalihokan sa enzyme sa succinate dehydrogenase. (7) Ang pagtaas sa lebel sa ROS mahimong makapugong sa kalihokan sa complex II. (8) Ang ubos nga kahusayan sa fatty acid oxidation nakatampo sa pagkunhod sa pagkaanaa sa substrate alang sa mitochondrial respiration. (9) Ang glycolysis gipauswag isip usa ka mekanismo sa kompensasyon alang sa pagkunhod sa produksiyon sa ATP sa oxidative phosphorylation. (10) Ang ATP synthase naglihok nga baliskad aron mapadayon ang ??m. Ang pula ug asul nagpaila sa upregulation ug downregulation, matag usa. Ang mga kahon nagpasabot sa pagkaanaa sa eksperimento nga ebidensya. Ang inset nagpakita sa mga hulagway sa mitochondria sa WT ug Nrf2-KO cortical astrocytes nga makita sa potentiometric fluorescent probe tetramethylrhodamine methyl ester (TMRM; 25 nM). Scale bar, 20 �m.
Kini nga mga kalainan sa m ug respirasyon sa taliwala sa mga genotypes makita sa rate sa paggamit sa mga substrate alang sa mitochondrial respiration. Paggamit sa substrates alang sa tricarboxylic acid (TCA) cycle (malate/pyruvate, nga sa baylo nagdugang sa produksyon sa complex I substrate NADH) o methyl succinate, usa ka substrate alang sa complex II, hinungdan sa usa ka stepwise nga pagtaas sa ??m sa duha WT ug Keap1-KD neurons, apan ang rate sa pagtaas mas taas sa Keap1-KD cells. Labaw sa tanan, ang mga porma sa tubag niini nga TCA cycle substrates lahi sa taliwala sa duha ka genotypes, diin ang paspas nga pagtaas sa m sa Keap1-KD nga mga selula sa substrate pagdugang gisundan sa usa ka dali nga pagtulo imbes sa usa ka patag, nga nagsugyot sa usa ka talagsaon nga. paspas nga pagkonsumo sa substrate. Kini nga mga nahibal-an nahiuyon sa labi ka ubos (sa 50-70%) nga lebel sa malate, pyruvate, ug succinate nga naobserbahan pagkahuman sa usa ka oras nga pulso sa [U-1C13] glucose sa Keap6-KO kumpara sa WT MEF mga selula [1]. Sa Nrf24-KO nga mga neuron, ang pyruvate lamang ang makahimo sa pagdugang sa m, samtang ang malate ug methyl succinate hinungdan sa malumo nga depolarization. Ang epekto sa Nrf2 sa produksyon sa mitochondrial substrate daw mao ang nag-unang mekanismo diin ang Nrf2 makaapekto sa mitochondrial function. Ang mitochondrial NADH redox index (ang balanse tali sa konsumo sa NADH pinaagi sa complex I ug produksyon sa NADPH sa TCA cycle) mas ubos sa Nrf2-KO nga mga selula kon itandi sa ilang WT nga mga katugbang, ug dugang pa, ang mga rate sa pagbag-o sa mga pool sa Ang NADH ug FADH2 human sa pagdili sa complex IV (pinaagi sa paggamit sa NaCN) mas hinay sa mutant cells.
Sa mitochondria nga nahimulag gikan sa utok sa murine ug atay, ang supplementation sa mga substrate alang sa complex I o alang sa complex II nagdugang sa rate sa konsumo sa oxygen nga mas kusog sa dihang ang Nrf2 gi-activate ug dili kaayo episyente kung ang Nrf2 nabalda [35]. Busa, ang malate nag-aghat sa mas taas nga rate sa konsumo sa oxygen sa Keap1-KD kumpara sa WT, apan ang epekto niini mas huyang sa Nrf2-KO mitochondria. Sa susama, sa presensya sa rotenone (kon ang complex I gipugngan), ang succinate nagpalihok sa pagkonsumo sa oksiheno sa mas dako nga gidak-on sa Keap1-KD kon itandi sa WT, samtang ang tubag sa Nrf2-KO mitochondria mikunhod. Dugang pa, ang Nrf2-KO nga nag-unang neuronal nga mga kultura ug mga ilaga mas sensitibo sa toxicity sa complex II inhibitors 3-nitropropionic acid ug malonate, samtang ang intrastriatal transplantation sa Nrf2-overexpressing astrocytes mao ang protective [48], [49]. Sa susama, ang Nrf2-KO nga mga ilaga mas sensitibo sa, samtang ang genetic o pharmacological activation sa Nrf2 adunay protective effects batok, neurotoxicity tungod sa complex I inhibitor 1-methyl-4-phenylpyridinium ion sa 1-methyl-4-phenyl-1,2,3,6, 49-tetrahydropyridine animal model sa Parkinson's disease [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61], [XNUMX].
Ang respiratory control ratio (RCR), ang ratio sa State 3 (ADP-stimulated) ngadto sa State 4 respiration (walay ADP present), mikunhod sa pagkawala sa Nrf2, apan ang RCR susama sa Keap1-KD ug WT mitochondria [35]. ]. Ingon nga ang RCR usa ka timailhan sa lebel sa pagdugtong sa kalihokan sa mitochondrial respiratory chain sa oxidative phosphorylation, kini nga pagpangita nagpakita nga ang mas taas nga rate sa respiration sa Keap1-KD mitochondria dili tungod sa uncoupling sa oxidative phosphorylation. Gisugyot pa niini nga ang oxidative phosphorylation mas episyente kung ang Nrf2 gi-aktibo. Ang mas taas nga rate sa respirasyon sa Keap1-KD mitochondria nahiuyon sa mas taas nga lebel sa produksiyon sa mitochondrial ROS [38] tungod kay ang mas taas nga rate sa respiration mahimong mosangpot sa pagtaas sa electron leak. Bisan pa, sa ilawom sa mga kondisyon sa tensiyon sa oxidative, ang pagtaas sa produksiyon sa ROS gikontra sa Nrf2-dependent transcriptional upregulation sa uncoupling protein 3 (UCP3), nga nagdugang sa proton conductance sa mitochondrial inner membrane ug sa ingon gipaubos ang produksiyon sa superoxide [62]. Bag-ohay lang, gipakita nga ang produkto sa lipid peroxidation 4-hydroxy-2-nonenal nagpataliwala sa Nrf2-dependent upregulation sa UCP3 sa cardiomyocytes; Kini mahimo nga labi ka hinungdanon alang sa pagpanalipod sa ilawom sa mga kondisyon sa stress sa oxidative sama sa panahon sa reperfusion sa ischemia [63].
Ang Nrf2 Makaapektar sa Episyente sa Oxidative Phosphorylation ug sa Synthesis sa ATP
Sa pag-uyon sa epekto sa Nrf2 sa respirasyon, sa utok ug atay mitochondria, ang kakulangan sa Nrf2 moresulta sa pagkunhod sa kahusayan sa oxidative phosphorylation (sumala sa gibanabana sa ratio sa ADP ngadto sa oxygen, nga gigamit alang sa ATP synthesis), samtang ang Nrf2 activation (Keap1). -KD) adunay kaatbang nga epekto [35]. Kung itandi sa WT, ang lebel sa ATP labi ka taas sa mga selyula nga adunay constitutive upregulation sa Nrf2 ug mas ubos kung ang Nrf2 natumba [64] o nabalda [35]. Dugang pa, ang paggamit sa mga inhibitor sa oxidative phosphorylation (oligomycin) o glycolysis (iodoacetic acid) nagpadayag nga ang Nrf2 nagbag-o sa paagi diin ang mga selula makahimo og ATP. Busa, sa WT neurons, oligomycin hinungdan sa usa ka bug-os nga drop sa ATP ug iodoacetic acid walay dugang nga epekto. Talagsaon, sa mga selula sa Nrf2-KO, ang oligomycin nagdugang sa lebel sa ATP, nga unya hinay-hinay, apan hingpit, nahurot sa iodoacetic acid, nga nagpakita nga sa pagkawala sa Nrf2, glycolysis, ug dili oxidative phosphorylation, mao ang nag-unang tinubdan sa produksyon sa ATP. Makapainteres, bisan pa sa dugang nga kahusayan sa oxidative phosphorylation sa Keap1-KD nga mga selula, ang pagdugang sa oligomycin nagresulta sa usa ka ~ 80% nga pagkunhod sa lebel sa ATP, ug ang iodoacetic acid hinungdan sa usa ka dugang nga ~ 20% nga pagkunhod. Busa, ang kakulangan sa Nrf2 o ang constitutive activation niini makapamenos sa kontribusyon sa oxidative phosphorylation ug makadugang sa kontribusyon sa glycolysis ngadto sa synthesis sa ATP. Kini nga epekto labi nga gipahayag kung wala ang Nrf2 ug nahiuyon sa pagsalig sa m sa presensya sa glucose sa medium [35] ug ang pagtaas sa lebel sa glycolytic intermediates (G-6-P, F-6-P , dihydroxyacetone phosphate, pyruvate, ug lactate) human sa knockdown sa Nrf2 [24].
Ang pagtaas sa lebel sa ATP human sa pagpugong sa F1F0-ATPase pinaagi sa oligomycin nagpakita nga kung wala ang Nrf2, ang F1F0-ATPase naglihok isip usa ka ATPase ug dili usa ka ATP synthase, ie, kini naglihok sa reverse. Ang ingon nga pagbag-o sa kalihokan lagmit nagpakita sa panginahanglan sa pagbomba sa mga proton tabok sa sulod nga mitochondrial membrane sa pagsulay nga mapadayon ang ??m, nga hinungdanon alang sa integridad sa pagpaandar niini nga organelle. Ang pagbag-o sa function sa F1F0-ATPase gipamatud-an usab sa naobserbahan nga mitochondrial depolarization sa oligomycin nga administrasyon sa Nrf2-KO nga mga selula, nga sukwahi sa hyperpolarization nga nahitabo sa ilang WT o Keap1-kulang nga mga katugbang [35]. Sa kinatibuk-an, daw ubos sa mga kondisyon sa Nrf2 deficiency ATP ang gihimo sa panguna sa glycolysis, ug kini nga ATP gigamit sa bahin sa F1F0-ATPase aron mapadayon ang ??m.
Ang Nrf2 Nagpauswag sa Mitochondrial Fatty Acid Oxidation
Ang epekto sa kakulangan sa Nrf2 sa ??m labi nga gipahayag kung ang mga selyula gilumlum sa medium nga walay glucose, ug ang ??m mao ang ~ 50% nga mas ubos sa Nrf2-KO kumpara sa mga selula sa WT [35]. Ubos sa mga kondisyon sa kakulangan sa glucose, ang mitochondrial fatty acid oxidation (FAO) usa ka mayor nga tighatag sa mga substrate alang sa respiration ug oxidative phosphorylation, nga nagsugyot nga ang Nrf2 mahimong makaapekto sa FAO. Sa tinuud, ang kahusayan sa FAO alang sa taas nga kadena (C16: 0) saturated fatty acid palmitic acid ug ang mubo nga kadena (C6: 0) hexanoic acid mas taas sa Keap1-KO MEFs ug nahimulag nga mitochondria sa kasingkasing ug atay kaysa sa ilang Ang mga katugbang sa WT, samtang kini mas ubos sa Nrf2-KO nga mga selula ug mitochondria [65]. Kini nga mga epekto usab adunay kalabotan sa mga tawo: sa tinuud, ang mga pagbag-o sa metaboliko nga nagpaila sa labi ka maayo nga panagsama sa FAO sa kalihokan sa siklo sa TCA gitaho nga mahitabo sa mga pagtuon sa interbensyon sa tawo nga adunay mga diyeta nga dato sa glucoraphanin, ang pasiuna sa klasikal nga Nrf2 activator sulforaphane [ 66].
Atol sa unang lakang sa mitochondrial FAO, ang pro-R hydrogen sa ?-carbon nga mga dahon isip usa ka hydride nga makapamenos sa FAD cofactor ngadto sa FADH2, nga sa baylo mobalhin sa mga electron ngadto sa ubiquinone (UbQ) sa respiratory chain, nga sa katapusan makatampo sa produksyon sa ATP . Samtang ang stimulation sa FAO pinaagi sa palmitoylcarnitine sa pagkawala sa glucose maoy hinungdan sa gipaabot nga pagtaas sa lebel sa ATP sa WT ug Keap1-KO nga mga selula, nga ang pagtaas sa ATP mas paspas sa Keap1-KO nga mga selula, ang parehas nga pagtambal walay mga pagbag-o sa ATP sa Nrf2-KO Mga MEF [65]. Gipakita niini nga eksperimento nga, kung wala ang Nrf2, ang FAO gipugngan, ug dugang pa, kini naglambigit sa pagsumpo sa FAO isip usa sa mga hinungdan sa ubos nga lebel sa ATP ubos sa mga kondisyon sa kakulangan sa Nrf2 [35], [64].
Ilabi na, ang mga selula sa 293 T sa tawo diin ang Nrf2 gipahilom adunay ubos nga ekspresyon sa CPT1 ug CPT2 [67], duha ka isoform sa carnitine palmitoyltransferase (CPT), ang rate-limiting enzyme sa mitochondrial FAO. Sa pag-uyon, ang lebel sa mRNA sa Cpt1 mas ubos sa mga atay sa Nrf2-KO kumpara sa WT nga mga ilaga [68]. Ang CPT nag-catalyze sa pagbalhin sa acyl group sa usa ka long-chain fatty acyl-CoA gikan sa coenzyme A ngadto sa l-carnitine ug sa ingon nagtugot sa import sa acylcarnitine gikan sa cytoplasm ngadto sa mitochondria. Bisan kung wala pa kini masusi hangtod karon, posible nga dugang sa mga epekto sa transkripsyon sa ekspresyon sa CPT1, ang Nrf2 mahimo usab nga makaapekto sa pag-obra niini nga enzyme pinaagi sa pagpugong sa lebel sa panguna nga allosteric inhibitor, malonyl-CoA. Kini tungod kay, pinaagi sa usa ka mekanismo nga sa pagkakaron dili klaro, ang Nrf2 nag-regulate sa negatibo nga ekspresyon sa stearoyl CoA desaturase (SCD) [69] ug citrate lyase (CL) [69], [70]. Katingad-an, ang pag-knockout o pagdili sa SCD modala ngadto sa dugang nga phosphorylation ug pagpaaktibo sa AMP-activated protein kinase (AMPK) [71], [72], [73], ug kini mahimong pangagpas nga, kung wala ang Nrf2, ang lebel sa SCD modaghan, sa baylo magpaubos sa kalihokan sa AMPK. Mahimo kini nga dugang nga madugangan sa pagkunhod sa lebel sa protina sa AMPK nga naobserbahan sa mga atay sa Nrf2-KO nga mga ilaga [68], usa ka pagpangita nga nahiuyon sa pagtaas sa lebel sa AMPK, nga gitaho sa mga atay sa Keap1-KD. mga ilaga [74]. Usa ka sangputanan sa pagkunhod sa kalihokan sa AMPK mao ang paghupay sa iyang inhibitory phosphorylation (sa Ser79) sa acetyl-CoA carboxylase (ACC) [75], nga mahimong dugang nga transcriptionally upregulated kung wala ang Nrf2 tungod kay kini gipaubos sa Nrf2 activation [70] ]. Ang taas nga kalihokan sa ACC, inubanan sa upregulated nga ekspresyon sa CL nga makadugang sa produksiyon sa acetyl-CoA, ang substrate alang sa ACC, mahimo nga sa katapusan madugangan ang lebel sa produkto sa ACC, malonyl-CoA. Ang taas nga lebel sa malonyl-CoA makapugong sa CPT, sa ingon makunhuran ang pagdala sa mga fatty acid ngadto sa mitochondria. Sa katapusan, ang Nrf2 positibo nga nag-regulate sa ekspresyon sa CD36 [76], usa ka translocase nga nag-import sa mga fatty acid sa plasma ug mitochondrial membranes. Busa, ang usa ka mekanismo diin ang Nrf2 mahimong makaapekto sa kahusayan sa mitochondrial FAO mao ang pag-regulate sa pag-import sa mga long-chain fatty acid ngadto sa mitochondria.
Gawas pa sa direktang regulasyon sa transkripsyon, ang Nrf2 mahimo usab nga mag-usab sa kahusayan sa mitochondrial FAO pinaagi sa mga epekto niini sa cellular redox metabolism. Mahimong kini ilabi na nga may kalabutan kung ang kalihokan sa Nrf2 ubos o wala, mga kondisyon nga nagbalhin sa cellular redox status ngadto sa oxidized state. Sa tinuud, daghang mga enzyme sa FAO ang giila nga sensitibo sa mga pagbag-o sa redox. Ang usa sa ingon nga enzyme mao ang taas kaayo nga kadena nga acyl-CoA dehydrogenase (VLCAD), nga nag-amot labaw sa 80% sa kalihokan sa dehydrogenation sa palmitoyl-CoA sa mga tisyu sa tawo [77]. Makaiikag, si Hurd et al. Gipakita sa [78] nga ang VLCAD adunay mga residu sa cysteine nga makabag-o sa ilang redox nga estado sa pagkaladlad sa nahilit nga mitochondria sa kasingkasing sa ilaga sa H2O2. Dugang pa, ang S-nitrosylation sa murine hepatic VLCAD sa Cys238 nagpauswag sa catalytic efficiency sa enzyme [79], ug lagmit nga ang oksihenasyon sa samang cysteine mahimong adunay kaatbang nga epekto, sa katapusan nagpaubos sa kahusayan sa mitochondrial FAO. Busa posible nga, bisan kung ang lebel sa ekspresyon sa VLCAD dili kaayo lahi sa WT, Nrf2-KO, o Keap1-KO MEFs [65], ang kalihokan sa enzyme sa VLCAD mahimong mubu kung wala ang Nrf2 tungod sa mas taas nga lebel. sa ROS.
Base sa tanan niini nga mga nahibal-an, mahimo nga isugyot nga (Fig. 3): kung wala ang Nrf2, ang lebel sa NADPH mas ubos tungod sa pagkunhod sa ekspresyon sa ME1, IDH1, G6PD, ug PGD. Ang lebel sa pagkunhod sa glutathione mas ubos usab tungod sa pagkunhod sa ekspresyon sa mga enzyme nga miapil sa biosynthesis ug pagbag-o niini ug ang mas ubos nga lebel sa NADPH nga gikinahanglan alang sa pagkakabig sa oxidized ngadto sa pagkunhod sa porma sa glutathione. Ang ubos nga ekspresyon sa ME1 makapakunhod sa pool sa pyruvate nga mosulod sa mitochondria, uban ang glycolysis nga nahimong mayor nga tinubdan sa pyruvate. Ang henerasyon sa NADH mas hinay, nga misangpot sa ningdaot nga kalihokan sa complex I ug nagdugang sa produksyon sa mitochondrial ROS. Ang pagkunhod sa FAD ngadto sa FADH2 mas hinay usab, labing menos sa usa ka bahin tungod sa dili kaayo episyente nga fatty acid oxidation, pagkompromiso sa electron flow gikan sa FADH2 ngadto sa UbQ ug ngadto sa complex III. Tungod kay ang UbQH2 usa ka activator sa succinate dehydrogenase [80], ang pagpahinay sa pagporma niini mahimong makapaubos sa kalihokan sa enzyme sa succinate dehydrogenase. Ang dugang nga lebel sa superoxide ug hydrogen peroxide mahimong makapugong sa komplikado nga II nga kalihokan sa dugang [81]. Ang ubos nga kaepektibo sa fatty acid oxidation nakatampo sa pagkunhod sa pagkaanaa sa substrate alang sa mitochondrial respiration ug paghimo sa ATP sa oxidative phosphorylation. Ingon usa ka mekanismo sa kompensasyon, ang glycolysis gipauswag. Ang ATP synthase naglihok nga balit-ad, isip usa ka ATPase, sa pagsulay sa pagpadayon sa ??m.
Nrf2 ug Mitochondrial Biogenesis
Gikataho nga, kon itandi sa WT, ang mga atay sa Nrf2-KO nga mga ilaga adunay ubos nga mitochondrial content (sumala sa gitino sa ratio sa mitochondrial ngadto sa nukleyar nga DNA); kini dugang nga pagkunhod sa usa ka 24-h nga pagpuasa sa WT ug Nrf2-KO nga mga ilaga; sa kasukwahi, bisan tuod walay kalainan sa WT ubos sa normal nga mga kondisyon sa pagpakaon, ang mitochondrial nga sulod sa mga ilaga nga adunay taas nga Nrf2 nga kalihokan wala maapektuhan sa pagpuasa [82]. Makapainteres, ang suplemento sa Nrf2 activator (R) -?-lipoic acid [83], [84], [85] nagpasiugda sa mitochondrial biogenesis sa 3T3-L1 adipocytes [86]. Duha ka klase sa nukleyar nga transcriptional regulators adunay mga kritikal nga papel sa mitochondrial biogenesis. Ang una nga klase mao ang transcription nga mga hinungdan, sama sa nukleyar nga respiratory factor11 ug 2, nga nagkontrol sa ekspresyon sa mga gene nga nag-encode sa mga subunit sa lima ka mga respiratory complex, mitochondrial translational nga mga sangkap, ug heme biosynthetic enzymes nga na-localize sa mitochondrial matrix [88]. Piantadosi et al. [89] nagpakita nga ang Nrf2-dependent transcriptional upregulation sa nuclear respiratory factor 1 nagpasiugda sa mitochondrial biogenesis ug nanalipod batok sa cytotoxicity sa cardiotoxic anthracycline chemotherapeutic agent doxorubicin. Sa kasukwahi, si Zhang et al. [82] nagtaho nga ang genetic activation sa Nrf2 wala makaapekto sa basal mRNA nga ekspresyon sa nuclear respiratory factor 1 sa murine liver.
Ang ikaduhang klase sa nuclear transcriptional regulators nga adunay kritikal nga mga function sa mitochondrial biogenesis mao ang transcriptional coactivators, sama sa peroxisome proliferator-activated receptor? coactivators (PGC)1? ug 1?, nga nakig-uban sa transcription nga mga hinungdan, ang basal transcriptional ug RNA-splicing machinery, ug histone-modifying enzymes [88], [90], [91]. Ang ekspresyon sa PGC1 nga pamilya sa mga coactivator naimpluwensyahan sa daghang mga signal sa kalikopan. Ang pagtambal sa mga fibroblast sa tawo nga adunay Nrf2 activator sulforaphane hinungdan sa pagtaas sa mitochondrial mass ug induction sa PGC1? ug PGC1? [92], bisan kung ang potensyal nga pagsalig sa Nrf2 wala masusi niini nga pagtuon. Bisan pa, ang mga ilaga sa diabetes diin ang Nrf2 gi-activate sa Keap1 gene hypomorphic knockdown (db/db:Keap1flox/?:Nrf2+/+) o nabalda (db/db:Keap1flox/?:Nrf2?/?) adunay ubos nga hepatic PGC1? lebel sa ekspresyon kay sa pagkontrolar sa mga mananap (db/db:Keap1flox/+:Nrf2+/+) [93]. Wala’y kalainan sa lebel sa mRNA alang sa PGC1? makita sa mga atay sa nondiabetic nga mga ilaga nga WT o Nrf2-KO, samtang kini nga lebel mas ubos sa Nrf2-overexpressing (Keap1-KD ug liver-specific Keap1-KO) nga mga hayop [82]. Talagsaon, ang usa ka 24-h nga pagpuasa nagdugang sa lebel sa PGC1? mRNA sa mga atay sa mga ilaga sa tanan nga mga genotypes, apan ang pagtaas mas dako sa mga atay sa Nrf2-KO kumpara sa WT o Nrf2-overexpressing nga mga ilaga. Kung itandi sa WT, Nrf2-KO nga mga ilaga nga nakasinati og septic infection o acute lung injury tungod sa impeksyon nagpakita sa attenuated transcriptional upregulation sa nuclear respiratory factor 1 ug PGC1? [94], [95]. Mag-uban, kini nga mga obserbasyon nagsugyot nga ang papel sa Nrf2 sa pagpadayon sa lebel sa nukleyar nga respiratory factor 1 ug PGC1? komplikado ug nahimong labing prominente ubos sa mga kondisyon sa stress.
Dugang sa pagpahayag sa mga gene nga nag-encode sa mga protina sa mitochondrial, ang mitochondrial biogenesis nagkinahanglan sa synthesis sa mga nucleotides. Ang genetic activation sa Nrf2 nagpalambo sa purine biosynthesis pinaagi sa pag-upregulate sa pentose phosphate nga agianan ug sa metabolismo sa folate ug glutamine, ilabi na sa paspas nga pagdaghan sa mga selula (Fig. 2) [24]. Ang pag-analisa sa transcriptome sa mutant Drosophila nga kulang alang sa mitochondrial serine / threonine protein kinase PTEN-induced putative kinase 1 (PINK1) nagpakita nga ang mitochondrial dysfunction mosangpot sa transcriptional upregulation sa mga gene nga naka-apekto sa metabolismo sa nucleotide [96], nga nagsugyot nga ang gipalambo nga nucleotide biosynthesis nagrepresentar sa usa ka mekanismo alang sa pagpanalipod batok sa neurotoxic nga mga sangputanan sa kakulangan sa PINK1. Ang Nrf2 nag-regulate sa ekspresyon sa phosphoribosyl pyrophosphate amidotransferase (PPAT), nga nag-catalyze sa pagsulod sa de novo purine nucleotide biosynthetic pathway, ug mitochondrial methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) (Fig. 2). Ang naulahi usa ka bifunctional enzyme nga adunay dehydrogenase ug cyclohydrolase nga mga kalihokan nga kritikal sa paghatag sa glycine ug formate isip mga tinubdan sa usa ka carbon units alang sa purine biosynthesis sa paspas nga pagtubo sa mga selula [97]. Busa lagmit nga ang pagpaaktibo sa Nrf2 mahimong mapanalipdan ug mahimong balihon ang mitochondrial dysfunction sa kakulangan sa PINK1. Sa tinuud, ang pagpaaktibo sa pharmacological sa Nrf2 pinaagi sa sulforaphane, o ang triterpenoid RTA-408, nagpahiuli sa m ug nanalipod sa mga selula nga kulang sa PINK1 batok sa pagkahilo sa dopamine [98]. Bisan kung ang nagpahiping mga mekanismo daw komplikado, magkauban, kini nga mga nahibal-an nagpakita nga ang kalihokan sa Nrf2 mahimong makaapekto sa mitochondrial biogenesis pinaagi sa pag-impluwensya sa lebel sa ekspresyon sa kritikal nga mga hinungdan sa transkripsyon ug mga coactivator, ingon man pinaagi sa pagpauswag sa biosynthesis sa nucleotide.
Nrf2 ug Mitochondrial Integrity
Bisan tuod ang direktang ebidensya dili kanunay anaa, adunay lig-on nga mga timailhan nga ang Nrf2 importante alang sa mitochondrial integrity, ilabi na ubos sa mga kondisyon sa oxidative stress. Ang mitochondria nga nahimulag gikan sa utok ug atay sa mga ilaga nga gihatag sa usa ka dosis sa Nrf2 activator sulforaphane dili makasugakod sa pag-abli sa mitochondrial permeability transition pore (mPTP) tungod sa oxidant tert-butylhydroperoxide [99], [100]. Ang mPTP, usa ka komplikado nga nagtugot sa mitochondrial sulod nga lamad nga mahimong permeable sa mga molekula nga adunay mga masa hangtod sa 1500 Da, bag-o lang giila nga naporma gikan sa mga dimer sa F0F1-ATP synthase [101]. Ang sulforaphane-mediated nga pagsukol sa pag-abli sa mPTP adunay kalabutan sa dugang nga mga depensa sa antioxidant, ug ang lebel sa mitochondrial GSH, glutathione peroxidase 1, malic enzyme 3, ug thioredoxin 2 tanan gi-upregulated sa mitochondrial fractions nga nahimulag gikan sa sulforaphane-treated nga mga hayop [100].
Ang kadaot sa protina sa mitochondrial ug pagkadaot sa respirasyon tungod sa electrophilic lipid peroxidation nga produkto nga 4-hydroxy-2-nonenal gipahinay sa mitochondria nga nahimulag gikan sa cerebral cortex sa sulforaphane-treated nga mga ilaga [102]. Sa rat renal epithelial cells ug sa kidney, ang sulforaphane nanalipod batok sa cisplatin- ug gentamicin-induced toxicity ug pagkawala sa ??m[103], [104]. Ang panalipod batok sa usa ka panel sa mga oxidant (superoxide, hydrogen peroxide, peroxynitrite) ug electrophiles (4-hydroxy-2-nonenal ug acrolein) ug usa ka pagtaas sa mitochondrial antioxidant defenses naobserbahan usab sa pagtambal sa rat aortic smooth muscle cells nga adunay sulforaphane [105]. ]. Sa usa ka modelo sa contrast-induced acute kidney injury, ang limb ischemic preconditioning bag-o lang gipakita nga adunay mga protective effect, lakip ang pagdili sa pag-abli sa mPTP ug mitochondrial swelling, pinaagi sa pagpaaktibo sa Nrf2 nga resulta sa pagdili sa GSK3? [106].
Ang mitophagy, ang proseso diin ang dysfunctional mitochondria gipili nga gilamoy sa mga autophagosome ug gihatud sa mga lysosome aron madaot ug ma-recycle sa selula, hinungdanon alang sa mitochondrial homeostasis [107], [108]. Samtang walay hinungdan nga relasyon tali sa Nrf2 ug mitophagy nga natukod, adunay ebidensya nga ang transcription factor mahimong importante sa pagkontrol sa kalidad sa mitochondrial pinaagi sa pagdula sa usa ka papel sa mitophagy. Mahimong kini ilabinang prominente ubos sa mga kondisyon sa oxidative stress. Busa, sa usa ka modelo sa sepsis, ang pagtaas sa lebel sa autophagosome marker MAP1 light chain 3-II (LC3-II) ug ang cargo protein p62 sa 24 h postinfection gipugngan sa Nrf2-KO kumpara sa WT nga mga ilaga [109] . Usa ka gamay nga molekula nga inducer sa mitophagy (gitawag nga p62-mediated mitophagy inducer, PMI) bag-o lang nadiskobrehan; kini nga 1,4-diphenyl-1,2,3-triazole compound orihinal nga gidisenyo isip Nrf2 activator nga makabalda sa interaksyon sa transcription factor sa Keap1 [110]. Susama sa mga selula diin ang Nrf2 gi-genetically upregulated (Keap1-KD o Keap1-KO), ang mga selula nga na-expose sa PMI adunay mas taas nga resting ??m. Importante, ang pagtaas sa mitochondrial LC3 localization nga naobserbahan human sa PMI nga pagtambal sa mga selula sa WT dili mahitabo sa Nrf2-KO nga mga selula, nga nagsugyot sa pagkalambigit sa Nrf2.
Ang katapusan, ang ultrastructural analysis sa mga seksyon sa atay nagpadayag sa presensya sa swollen mitochondria nga adunay pagkunhod sa crista ug disrupted membranes sa hepatocytes sa Nrf2-KO, apan dili WT, mga ilaga nga gipakaon sa taas nga tambok nga pagkaon alang sa 24 nga mga semana; ilabi na, kini nga mga atay nagpakita sa tin-aw nga ebidensya sa oxidative stress ug panghubag [68]. Mahimong makahinapos nga ang Nrf2 adunay usa ka kritikal nga papel sa pagpadayon sa integridad sa mitochondrial ubos sa mga kondisyon sa oxidative ug makapahubag nga stress.
Sulforaphane ug ang mga Epekto Niini sa Kanser, Pagka-mortal, Pagkatigulang, Utok ug Panggawi, Sakit sa Kasingkasing ug uban pa
Ang Isothiocyanates mao ang pipila sa labing hinungdanon nga mga compound sa tanum nga makuha nimo sa imong pagkaon. Niini nga video gihimo nako ang labing komprehensibo nga kaso alang kanila nga nahimo sukad. Mubo nga gidugayon sa pagtagad? Laktaw sa imong paborito nga hilisgutan pinaagi sa pag-klik sa usa sa mga punto sa oras sa ubos. Full timeline sa ubos.
Pangunang mga seksyon:
00:01:14 – Kanser ug mortalidad
00:19:04 - Pagkatigulang
00:26:30 - Utok ug pamatasan
00:38:06 - Katapusan nga recap
00:40:27 - Dosis
Tibuok timeline:
00:00:34 - Pagpaila sa sulforaphane, usa ka mayor nga focus sa video.
00:01:14 - Pagkonsumo sa cruciferous nga utanon ug pagkunhod sa tanan nga hinungdan sa pagkamatay.
00:02:12 - Risgo sa kanser sa prostate.
00:02:23 - Risgo sa kanser sa pantog.
00:02:34 - Ang kanser sa baga sa mga nanigarilyo peligro.
00:02:48 - Risgo sa kanser sa suso.
00:03:13 - Hypothetical: unsa man kung ikaw adunay kanser? (interbensyonal)
00:03:35 - Katuohan nga mekanismo nga nagmaneho sa kanser ug mortalidad nga asosasyon nga datos.
00:04:38 - Sulforaphane ug kanser.
00:05:32 - Ang ebidensya sa hayop nga nagpakita sa kusog nga epekto sa broccoli sprout extract sa pag-uswag sa tumor sa pantog sa mga ilaga.
00:06:06 - Epekto sa direktang suplemento sa sulforaphane sa mga pasyente sa kanser sa prostate.
00:07:09 - Bioaccumulation sa isothiocyanate metabolites sa aktwal nga tisyu sa dughan.
00:08:32 - Pagpugong sa mga stem cell sa kanser sa suso.
00:08:53 - Leksyon sa kasaysayan: ang mga brassicas gitukod nga adunay mga kabtangan sa kahimsog bisan sa karaang Roma.
00:09:16 - Ang abilidad sa Sulforaphane sa pagpalambo sa carcinogen excretion (benzene, acrolein).
00:10:10 - Sa unsang paagi ang pagpaaktibo sa NRF2 makapauswag sa carcinogen excretion pinaagi sa glutathione-S-conjugates.
00:10:34 - Ang Brussels sprouts nagdugang sa glutathione-S-transferase ug makapamenos sa kadaot sa DNA.
00:11:20 - Ang broccoli sprout nga ilimnon nagdugang sa benzene excretion sa 61%.
00:13:31 - Ang homogenate nga pagtubo sa broccoli nagdugang antioxidant enzymes sa taas nga agianan sa hangin.
00:15:45 - Pagkonsumo sa cruciferous nga utanon ug pagkamatay sa sakit sa kasingkasing.
00:16:55 - Ang broccoli sprout powder nagpauswag sa mga lipid sa dugo ug sa kinatibuk-ang risgo sa sakit sa kasingkasing sa type 2 nga mga diabetes.
00:19:04 - Pagsugod sa seksyon sa pagkatigulang.
00:19:21 - Sulforaphane-enriched nga pagkaon nagpauswag sa kinabuhi sa mga bakukang gikan sa 15 ngadto sa 30% (sa pipila ka mga kondisyon).
00:20:34 - Ang kamahinungdanon sa ubos nga panghubag alang sa taas nga kinabuhi.
00:22:05 - Ang cruciferous nga mga utanon ug broccoli sprout powder daw makapakunhod sa nagkalainlain nga mga marker sa panghubag sa mga tawo.
00:23:40 - Mid-video recap: kanser, mga seksyon sa pagkatigulang
00:24:14 - Ang mga pagtuon sa mouse nagsugyot nga ang sulforaphane mahimong makapauswag sa adaptive immune function sa pagkatigulang.
00:25:18 - Ang Sulforaphane nagpauswag sa pagtubo sa buhok sa usa ka modelo sa mouse sa pagkalbo. Hulagway sa 00:26:10.
00:26:30 - Pagsugod sa seksyon sa utok ug pamatasan.
00:27:18 - Epekto sa broccoli sprout extract sa autism.
00:27:48 - Epekto sa glucoraphanin sa schizophrenia.
00:28:17 - Pagsugod sa diskusyon sa depresyon (katuohan nga mekanismo ug pagtuon).
00:31:21 - Ang pagtuon sa mouse gamit ang 10 ka lain-laing mga modelo sa stress-induced depression nagpakita sa sulforaphane nga parehas nga epektibo sama sa fluoxetine (prozac).
00:32:00 - Gipakita sa pagtuon nga ang direkta nga pag-inom sa glucoraphanin sa mga ilaga parehas nga epektibo sa pagpugong sa depresyon gikan sa sosyal nga kapildihan nga modelo sa stress.
00:33:01 - Pagsugod sa seksyon sa neurodegeneration.
00:33:30 - Sulforaphane ug Alzheimer's disease.
00:33:44 - Sulforaphane ug Parkinson's disease.
00:33:51 - Sulforaphane ug sakit sa Hungtington.
00:34:13 - Ang Sulforaphane nagdugang sa mga protina sa heat shock.
00:34:43 - Pagsugod sa seksyon sa traumatic brain injury.
00:35:01 - Sulforaphane gi-injected dayon human ang TBI nagpalambo sa memorya (pagtuon sa mouse).
00:35:55 - Sulforaphane ug neuronal plasticity.
00:36:32 - Ang Sulforaphane nagpauswag sa pagkat-on sa modelo sa type II diabetes sa mga ilaga.
00:37:19 - Sulforaphane ug duchenne muscular dystrophy.
00:37:44 - Pagpugong sa Myostatin sa mga selula sa satellite sa kalamnan (in vitro).
00:38:06 - Late-video recap: mortalidad ug kanser, kadaot sa DNA, oxidative stress ug panghubag, benzene excretion, sakit sa cardiovascular, type II diabetes, mga epekto sa utok (depresyon, autism, schizophrenia, neurodegeneration), NRF2 nga agianan.
00:40:27 - Mga hunahuna sa paghunahuna sa usa ka dosis sa broccoli sprouts o sulforaphane.
00:41:01 - Mga anekdota sa pagtubo sa balay.
00:43:14 - Sa temperatura sa pagluto ug kalihokan sa sulforaphane.
00:43:45 - Ang pagkakabig sa bakterya sa gut sa sulforaphane gikan sa glucoraphanin.
00:44:24 - Ang mga suplemento mas maayo kung gihiusa sa aktibo nga myrosinase gikan sa mga utanon.
00:44:56 - Mga teknik sa pagluto ug cruciferous nga mga utanon.
00:46:06 - Isothiocyanates isip goitrogens.
Ang Nrf2 usa ka transcription factor nga adunay hinungdanon nga papel sa cellular antioxidant defense system sa lawas sa tawo. Ang antioxidant responsive nga elemento, o ARE, usa ka mekanismo sa regulasyon sa mga gene. Daghang mga pagtuon sa panukiduki ang nagpakita nga ang Nrf2, o NF-E2-related factor 2, nag-regulate sa usa ka halapad nga klase sa ARE-driven nga mga gene sa daghang mga matang sa mga selula. Ang Nrf2 nakit-an usab nga adunay hinungdanon nga papel sa pagpanalipod sa cellular ug anti-carcinogenicity, nga nagpakita nga ang Nrf2 mahimo’g usa ka epektibo nga pagtambal sa pagdumala sa mga sakit nga neurodegenerative ug mga kanser nga gituohan nga gipahinabo sa oxidative stress. Dr. Alex Jimenez DC, CCST Insight
Panapos nga Pulong
Bisan kung daghang mga pangutana ang nagpabilin nga bukas, ang magamit nga ebidensya sa eksperimento tin-aw nga nagpakita nga ang Nrf2 usa ka hinungdanon nga magdudula sa pagpadayon sa mitochondrial homeostasis ug integridad sa istruktura. Kini nga tahas nahimong labi ka kritikal ubos sa mga kondisyon sa oxidative, electrophilic, ug makapahubag nga stress kung ang abilidad sa pag-upregulate sa Nrf2-mediated cytoprotective nga mga tubag makaimpluwensya sa kinatibuk-ang kahimsog ug survival sa cell ug sa organismo. Ang papel sa Nrf2 sa mitochondrial function nagrepresentar sa laing layer sa halapad nga cytoprotective nga mga mekanismo nga gi-orkestra niini nga transcription factor. Sama sa daghang mga kondisyon sa pathological sa tawo nga adunay oxidative stress, panghubag, ug mitochondrial dysfunction ingon hinungdanon nga sangkap sa ilang pathogenesis, ang pagpaaktibo sa pharmacological sa Nrf2 adunay saad alang sa paglikay ug pagtambal sa sakit. Ang komprehensibo nga pagsabut sa tukma nga mga mekanismo diin ang Nrf2 nakaapekto sa mitochondrial function hinungdanon alang sa makatarunganon nga laraw sa umaabot nga mga pagsulay sa klinika ug mahimo’g magtanyag bag-ong mga biomarker alang sa pagmonitor sa pagkaepektibo sa pagtambal.
Ang katuyoan sa artikulo sa ibabaw mao ang paghisgot ug pagpasundayag sa mitumaw nga papel sa Nrf2 sa mitochondrial function. Nrf2, o nuclear factor erythroid 2-related nga butang, mao ang usa ka emerging regulator sa cellular resistensya sa oxidants nga makatampo sa oxidative stress, makaapekto sa cellular function ug modala ngadto sa kalamboan sa toxicity, laygay nga sakit, ug bisan sa kanser. Samtang ang produksiyon sa mga oxidant sa lawas sa tawo mahimong magsilbi sa lain-laing mga katuyoan, lakip ang cell division, panghubag, immune function, autophagy, ug pagtubag sa stress, hinungdanon nga kontrolon ang ilang sobra nga produksiyon aron malikayan ang mga isyu sa kahimsog. Ang sakup sa among kasayuran limitado sa mga isyu sa kahimsog sa chiropractic ug spinal. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Dugang nga Hisgutan sa Hilisgutan:�Acute Back Pain
Sakit sa likod�usa sa labing kaylap nga hinungdan sa pagkabaldado ug wala’y trabaho nga mga adlaw sa tibuuk kalibutan. Ang sakit sa bukobuko hinungdan sa ikaduha nga kasagarang hinungdan sa pagbisita sa opisina sa doktor, nga mas daghan kaysa mga impeksyon sa taas nga respiratoryo. Gibana-bana nga 80 porsyento sa populasyon ang makasinati og sakit sa likod labing menos kausa sa tibuok nilang kinabuhi. Ang dugokan maoy usa ka komplikadong gambalay nga gilangkoban sa mga bukog, lutahan, ligaments, ug kaunuran, ug uban pang humok nga mga tisyu. Tungod niini, mga kadaot ug/o nagkagrabe nga mga kondisyon, sama sa�herniated discs, mahimong mosangpot sa mga sintomas sa sakit sa likod. Ang mga samad sa sports o mga aksidente sa awto sa kasagaran mao ang labing kanunay nga hinungdan sa sakit sa bukobuko, bisan pa, usahay ang pinakasimple nga mga paglihok mahimong adunay sakit nga mga sangputanan. Maayo na lang, ang alternatibong mga opsyon sa pagtambal, sama sa pag-atiman sa chiropractic, makatabang sa pagpagaan sa sakit sa likod pinaagi sa paggamit sa mga pag-adjust sa taludtod ug mga pagmaniobra sa manwal, nga sa katapusan makapauswag sa paghupay sa kasakit. �
Nrf2 nagsuporta sa pagpaaktibo sa usa ka grupo sa antioxidant ug detoxifying enzymes ug mga gene nga nanalipod sa lawas sa tawo gikan sa mga epekto sa mga isyu sa panglawas nga nalangkit sa dugang nga lebel sa oxidative stress, sama sa Alzheimer's disease. Ang nagkalainlain nga natural nga mga substansiya gipakita aron ma-activate ang Nrf2 nga agianan, nga makatabang sa pagdumala sa mga sintomas sa neurodegenerative nga mga sakit. Ang katuyoan sa artikulo sa ubos mao ang paghisgot sa hinungdanon nga papel sa Nrf2 tungod sa laygay nga panghubag.
abstract
Ang panghubag mao ang labing kasagaran nga bahin sa daghang mga sakit ug komplikasyon, samtang nagdula mga kritikal nga papel sa carcinogenesis. Daghang mga pagtuon ang nagpakita nga ang Nrf2 nakatampo sa anti-inflammatory nga proseso pinaagi sa pag-orkestra sa pagrekrut sa mga makapahubag nga mga selula ug pag-regulate sa ekspresyon sa gene pinaagi sa antioxidant response element (ARE). Ang Keap1 (Kelch-like ECH-associated protein)/Nrf2 (NF-E2 p45-related factor 2)/ARE signaling pathway nag-una nga nag-regulate sa anti-inflammatory gene expression ug nagpugong sa pag-uswag sa panghubag. Busa, ang pag-ila sa bag-ong Nrf2-dependent nga anti-inflammatory phytochemicals nahimong importanteng punto sa pagdiskobre sa droga. Niini nga pagrepaso, atong hisgutan ang mga miyembro sa Keap1 / Nrf2 / ARE signal pathway ug ang downstream genes niini, ang mga epekto niini nga agianan sa mga modelo sa mananap sa mga sakit nga makapahubag, ug crosstalk sa NF-?B nga agianan. Dugang pa, gihisgutan usab namon ang bahin sa regulasyon sa NLRP3 inflammasome sa Nrf2. Gawas pa niini, gi-summarize namo ang kasamtangan nga senaryo sa pagpalambo sa mga anti-inflammatory phytochemicals ug uban pa nga nagpataliwala sa Nrf2 / ARE signaling pathway.
Ang panghubag usa ka komplikado nga proseso nga mahitabo kung ang mga tisyu nataptan o nasamdan sa makadaot nga mga stimuli sama sa mga pathogen, kadaot, o mga irritant. Ang mga immune cell, mga ugat sa dugo, ug mga molekular nga tigpataliwala nalangkit sa kini nga pagpanalipod nga tubag [1]. Ang panghubag usa usab ka patolohiya nga panghitabo nga nalangkit sa lain-laing mga estado sa sakit nga gipahinabo sa panguna sa pisikal, kemikal, biolohikal, ug sikolohikal nga mga hinungdan. Ang tumong sa panghubag mao ang paglimite ug pagwagtang sa mga hinungdan sa kadaot sa selula, paghawan ug/o pagsuhop sa mga necrotic nga mga selula ug tisyu, ug pagsugod sa pag-ayo sa tisyu. Duha ka lahi nga porma sa panghubag ang gipalahi: acute ug chronic. Ang mahait nga panghubag nagpugong sa kaugalingon ug mapuslanon sa host, apan ang dugay nga laygay nga panghubag usa ka sagad nga bahin sa daghang mga sakit ug komplikasyon. Direkta nga paglusot sa daghang mononuclear immune cells sama sa monocytes, macrophage, lymphocytes, ug plasma cells, ingon man ang pagprodyus sa makapahubag nga mga cytokine, mosangpot sa chronic inflammation. Giila nga ang laygay nga panghubag adunay hinungdanon nga papel sa carcinogenesis [2]. Sa kinatibuk-an, ang mga pro- ug anti-inflammatory signaling pathways nakig-interact sa normal nga proseso sa pagpanghubag.
Sa proseso sa panghubag sa pathological, ang mga mast cell, monocytes, macrophage, lymphocytes, ug uban pang mga immune cell una nga gi-aktibo. Dayon ang mga selula gi-recruit sa dapit sa kadaot, nga miresulta sa pagmugna sa reactive oxygen species (ROS) nga makadaot sa macromolecules lakip na ang DNA. Sa samang higayon, kini nga mga selula sa pagpanghubag naghimo usab og daghang mga tigpataliwala sa panghubag sama sa mga cytokine, chemokines, ug prostaglandin. Kini nga mga tigpataliwala dugang nga nagrekrut sa mga macrophage sa mga lokal nga lugar sa panghubag ug direkta nga gi-aktibo ang daghang mga signal transduction cascades ug mga hinungdan sa transkripsyon nga may kalabotan sa panghubag. Ang NF-?B (nuclear factor kappa B), MAPK (mitogen-activated protein kinase), ug JAK (janus kinase) -STAT (signal transducers ug activators of transcription) signaling pathways nalangkit sa pagpalambo sa classical pathway of inflammation [3], [4], [5]. Gipadayag sa nangaging mga pagtuon nga ang transcription factor Nrf2 (NF-E2 p45-related factor 2) nag-regulate sa ekspresyon sa phase II detoxifying enzymes lakip na ang NADPH, NAD (P) H quinone oxidoreductase 1, glutathione peroxidase, ferritin, heme oxygenase-1 (H O -1), ug antioxidant nga mga gene nga nanalipod sa mga selula gikan sa lainlaing mga kadaot pinaagi sa ilang mga anti-inflammatory nga epekto, sa ingon nakaimpluwensya sa dagan sa sakit [6], [7], [8].
Sa pagkonsiderar niining talagsaon nga mga nahibal-an, ang pag-uswag sa gipunting nga mga tambal nga terapyutik alang sa mga sakit nga makapahubag pinaagi sa mga agianan sa pagsenyas nakadani sa daghang interes sa bag-ohay nga mga tuig. Niini nga pagrepaso, gi-summarize namo ang panukiduki sa Keap1 (Kelch-like ECH associated protein)/Nrf2 (NF-E2 p45-related factor 2)/ARE (antioxidant response element) signaling pathway sa panghubag.
Istruktura ug Regulasyon sa Nrf2
Keap1-Dependent Nrf2 Regulasyon
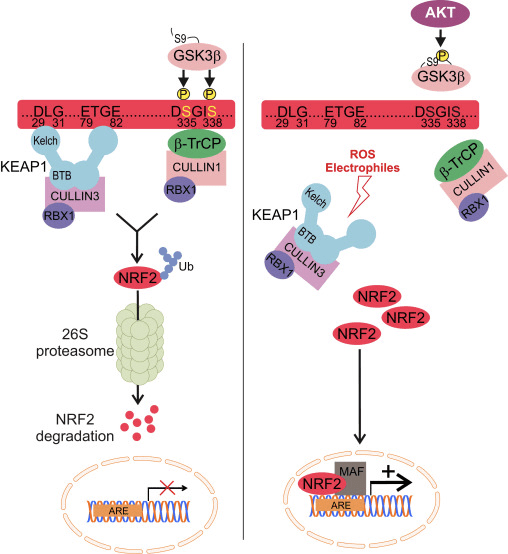
Ang Nrf2 iya sa Cap �n� Collar (CNC) subfamily ug naglangkob sa pito ka functional domains, Neh (Nrf2-ECH homology) 1 ngadto sa Neh7 [9], [10]. Ang Neh1 usa ka domain sa CNC-bZIP nga nagtugot sa Nrf2 nga heterodimerize sa gamay nga musculoaponeurotic fibrosarcoma (Maf) nga protina, DNA, ug uban pang mga kasosyo sa transkripsyon ingon man pagporma sa usa ka nukleyar nga komplikado nga adunay ubiquitin-conjugating enzyme UbcM2 [11], [12]. Ang Neh2 naglangkob sa duha ka importante nga motif nga nailhan nga DLG ug ETGE, nga gikinahanglan alang sa interaksyon tali sa Nrf2 ug sa negatibo nga regulator niini nga Keap1 [13], [14].
Ang Keap1 usa ka substrate adapter alang sa cullin-based E3 ubiquitin ligase, nga nagpugong sa transcriptional nga kalihokan sa Nrf2 pinaagi sa ubiquitination ug proteasomal degradation ubos sa normal nga kondisyon [15], [16], [17]. Ang KELCH domain sa Keap1 homodimer nagbugkos sa DLG ug ETGE motifs sa Nrf2-Neh2 domain sa cytosol, diin ang ETGE naglihok isip bisagra nga adunay mas taas nga kalambigitan ug ang DLG naglihok isip usa ka latch [18]. Ubos sa oxidative stress o sa pagkaladlad sa Nrf2 activators, Nrf2 dissociates gikan sa Keap1 binding tungod sa thiol modification sa Keap1 cysteine residues nga sa katapusan makapugong sa Nrf2 ubiquitination ug proteasomal degradation [19]. Unya Nrf2 translocates ngadto sa nucleus, heterodimerizes uban sa gagmay nga Maf protina, ug transactivates usa ka ARE battery sa mga gene (Fig. 1A). Ang carboxy-terminal sa Neh3 naglihok isip usa ka transactivation domain pinaagi sa pagpakig-uban sa transcription co-activator nga nailhan nga CHD6 (chromo-ATPase/helicase DNA binding protein) [20]. Ang Neh4 ug Neh5 naglihok usab isip mga domain sa transactivation, apan nagbugkos sa laing transcriptional co-activator nga nailhan nga CBP (cAMP-response-element-binding protein-binding protein) [21]. Dugang pa, ang Neh4 ug Neh5 nakig-uban sa nukleyar nga cofactor nga RAC3 / AIB1 / SRC-3, nga misangpot sa gipaayo nga Nrf2-target nga ARE nga ekspresyon sa gene [22]. Ang Neh5 adunay redox-sensitive nuclear-export signal nga hinungdanon alang sa regulasyon ug cellular localization sa Nrf2 [23].
Figure 1 Keap1-depende ug -independente nga regulasyon sa Nrf2. (A) Ubos sa basal nga kondisyon, ang Nrf2 gisunod sa Keap1 pinaagi sa duha ka motif niini (ETGE ug DLG) nga mosangpot sa CUL3-mediated ubiquitination nga gisundan sa proteasome degradation. Ubos sa stress sa oxidative, ang Nrf2 nagbulag gikan sa Keap1, nag-translocate sa nucleus ug gi-aktibo ang ARE-gene nga baterya. (B) GSK3 phosphorylates Nrf2 ug kini nagpadali sa pag-ila sa Nrf2 pinaagi sa ?-TrCP alang sa CUL1-mediated ubiquitination ug sa sunod nga proteasome degradation. (C) p62 gisequester sa Keap1, nga mitultol sa autophagic degradation niini, ang pagpalingkawas sa Nrf2, ug nadugangan ang Nrf2 signaling.
Keap1-Independente nga Regulasyon sa Nrf2
Ang nag-uswag nga ebidensya nagpadayag usa ka bag-ong mekanismo sa regulasyon sa Nrf2 nga independente sa Keap1. Ang dato sa serine nga Neh6 nga domain sa Nrf2 adunay hinungdanon nga papel sa kini nga regulasyon pinaagi sa pagbugkos sa duha nga mga motif niini (DSGIS ug DSAPGS) sa ?-transducin repeat-containing protein (?-TrCP) [24]. Ang ?-TrCP usa ka substrate receptor alang sa Skp1�Cul1�Rbx1/Roc1 ubiquitin ligase complex nga nagpunting sa Nrf2 alang sa ubiquitination ug proteasomal degradation. Glycogen synthase kinase-3 mao ang usa ka importante nga protina nga nalambigit sa Keap1-independente Nrf2 stabilization ug regulasyon; kini phosphorylates Nrf2 sa Neh6 domain aron mapadali ang pag-ila sa Nrf2 pinaagi sa ?-TrCP ug sunod-sunod nga protina degradation [25] (Fig. 1B).
Ang ubang mga Nrf2 Regulator
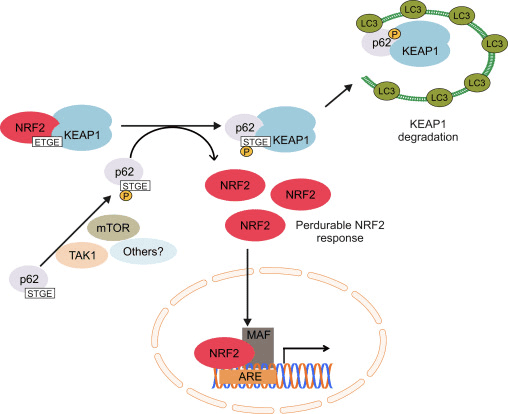
Ang laing linya sa ebidensya nagpadayag sa usa ka non-canonical pathway sa p62-dependent Nrf2 activation diin ang p62 sequesters Keap1 ngadto sa autophagic degradation nga sa katapusan modala ngadto sa stabilization sa Nrf2 ug ang transactivation sa Nrf2-dependent genes [26], [27], [ 28], [29] (Fig. 1C).
Ang pagtipon sa ebidensya nagsugyot nga daghang mga miRNA ang adunay hinungdanon nga papel sa regulasyon sa kalihokan sa Nrf2 [30]. Sangokoya et al. Gipakita sa [31] nga ang miR-144 direkta nga nagpaubos sa kalihokan sa Nrf2 sa lymphoblast K562 cell line, panguna nga human erythroid progenitor cells, ug sickle-cell disease reticulocytes. Ang laing makapaikag nga pagtuon sa mga selula sa epithelial sa dughan sa tawo nagpakita nga ang miR-28 nagpugong sa Nrf2 pinaagi sa mekanismo nga independente sa Keap1 [32]. Sa susama, ang mga miRNA sama sa miR-153, miR-27a, miR-142-5p, ug miR144 nagpaubos sa Nrf2 nga ekspresyon sa neuronal SH-SY5Y cell line [33]. Singh ug uban pa. Gipakita sa [34] nga ang ectopic nga ekspresyon sa miR-93 nagpamenos sa ekspresyon sa Nrf2-regulated nga mga gene sa usa ka 17? -estradiol (E2) -induced rat model sa mammary carcinogenesis.
Ang usa ka bag-o nga nadiskobrehan gikan sa among lab nagpaila sa usa ka endogenous inhibitor sa Nrf2 nga nailhan nga retinoic X receptor alpha (RXR?). RXR? usa ka nukleyar nga receptor, nakig-uban sa Neh7 domain sa Nrf2 (amino-acid residues 209-316) pinaagi sa DNA-binding domain (DBD), ug espesipikong nagpugong sa kalihokan sa Nrf2 sa nucleus. Dugang pa, ang ubang mga nukleyar nga receptor sama sa peroxisome proliferator-activated receptor-?, ER?, estrogen-related receptor-?, ug glucocorticoid receptors usab gitaho nga endogenous inhibitors sa Nrf2 activity [9], [10].
Anti-Inflammatory nga Papel sa Nrf2 / HO-1 Axis
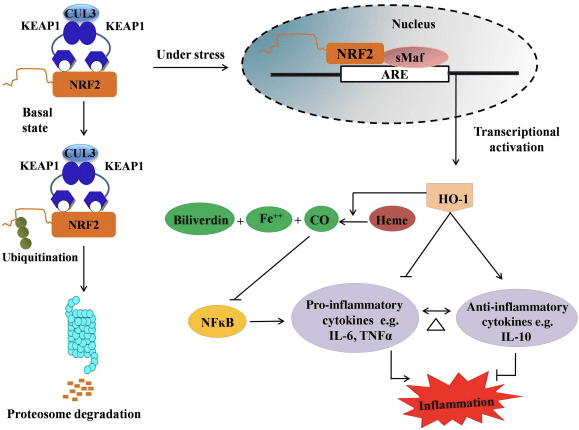
Ang HO-1 mao ang inducible isoform ug rate-limiting enzyme nga nag-catalyze sa pagkadaot sa heme ngadto sa carbon monoxide (CO) ug libreng iron, ug biliverdin ngadto sa bilirubin. Enzymatic degradation sa pro-inflammatory free heme ingon man ang produksyon sa mga anti-inflammatory compounds sama sa CO ug bilirubin adunay dagkong papel sa pagmintinar sa protective effects sa HO-1 (Fig. 2).
Figure 2 Overview sa Nrf2/HO-1 nga agianan. Ubos sa basal nga mga kondisyon, ang Nrf2 nagbugkos sa iyang repressor Keap1 nga mosangpot sa ubiquitination nga gisundan sa proteasome degradation. Atol sa oxidative stress, ang libre nga Nrf2 nag-translocate sa nucleus, diin kini nagdimerize sa mga miyembro sa gamay nga pamilya sa Maf ug nagbugkos sa ARE nga mga gene sama sa HO-1. Ang na-upregulated nga HO-1 nag-catalyze sa heme ngadto sa CO, bilirubin, ug libre nga iron. Ang CO naglihok isip usa ka tigpugong sa NF-?B nga agianan nga mosangpot sa pagkunhod sa ekspresyon sa pro-inflammatory cytokines, samtang ang bilirubin naglihok usab isip antioxidant. Dugang pa, ang HO-1 direktang nagpugong sa proinflammatory cytokines ingon man usab sa pagpaaktibo sa anti-inflammatory cytokines, sa ingon mosangpot sa pagbalanse sa proseso sa panghubag.
Ang Nrf2 nag-aghat sa HO-1 nga gene pinaagi sa pagdugang sa mRNA ug protina nga ekspresyon ug kini usa sa classic nga Nrf2 regulated gene nga kaylap nga gigamit sa daghang mga in vitro ug in vivo nga mga pagtuon. Gipakita sa daghang mga pagtuon nga ang HO-1 ug ang mga metabolite niini adunay hinungdanon nga mga epekto nga anti-inflammatory nga gipataliwala sa Nrf2. Ang pagtaas sa HO-1 nga ekspresyon nga gipataliwad-an sa gi-activate nga Nrf2 nagdala ngadto sa pagdili sa NF?B signaling nga resulta sa pagkunhod sa intestinal mucosal injury ug tight-junction dysfunction sa laki nga Sprague-Dawley rat liver transplantation model [35]. Ang upregulation sa Nrf2-dependent nga HO-1 nga ekspresyon mahimong manalipod sa mouse nga nakuha sa C2C12 myoblasts gikan sa H2O2 cytotoxicity [36]. Ang Nrf2-dependent nga HO-1 adunay epekto sa lipopolysaccharide (LPS) -mediated inflammatory responses sa RAW264.7- o mouse peritoneal macrophage-derived foam cell macrophage. Ang kalihokan sa Nrf2 desensitized foam cell macrophage phenotype ug mapugngan ang dili kasarangan nga paghubag sa mga macrophage, kini adunay hinungdanon nga papel sa pag-uswag sa atherosclerosis [37]. Ang Nrf2 / HO-1 axis nakaapekto sa LPS nga gipahinabo sa mouse BV2 microglial cells ug mouse hippocampal HT22 cells, nga adunay epekto sa neuroinflammation. Upregulation sa HO-1 nga ekspresyon pinaagi sa Nrf2 nga agianan sa mouse BV2 microglial cells nga nagdepensa sa cell death sa mouse hippocampal HT22 cells [38]. Dugang pa, ang cobalt-based hybrid molecules (HYCOs) nga naghiusa sa usa ka Nrf2 inducer nga adunay releaser sa carbon monoxide (CO) nagdugang sa Nrf2 / HO-1 nga ekspresyon, nagpalingkawas sa CO ug naggamit sa anti-inflammatory nga kalihokan sa vitro. Ang mga HYCO usab nag-regulate sa tisyu HO-1 ug naghatud sa CO sa dugo pagkahuman sa administrasyon sa vivo, nagsuporta sa ilang potensyal nga paggamit batok sa mga kondisyon nga makapahubag [39]. Ang Nrf2 / HO-1 upregulation makapakunhod sa panghubag pinaagi sa pagdugang sa efferocytic nga kalihokan sa murine macrophage nga gitambalan sa taurine chloramines [40]. Sa kinatibuk-an, ang gipatin-aw sa ibabaw nga mga eksperimento nga mga modelo nagpadayag nga ang Nrf2 / HO-1 axis adunay dakong papel sa anti-inflammatory function, nga nagsugyot nga ang Nrf2 usa ka therapeutic target sa mga sakit nga nalangkit sa panghubag.
Dugang pa, ang mga byproduct sa HO-1 sama sa CO, bilirubin, naglihok isip usa ka gamhanan nga antioxidant atol sa oxidative stress ug cell damage [41], [42]; gipugngan niini ang autoimmune encephalomyelitis ug hepatitis [43], [44]; ug kini nanalipod sa mga ilaga ug mga ilaga batok sa endotoxic shock pinaagi sa pagpugong sa henerasyon sa iNOS ug NO [45], [46], [47]. Dugang pa, ang Bilirubin nagpamenos sa endothelial activation ug dysfunction [48]. Makapainteres, ang bilirubin nagpamenos sa transmigration sa endothelial leukocytes pinaagi sa adhesion molecule-1 [49]. Kini nga mga piho nga mga pakisayran nga nagpaila dili lamang ang HO-1 naglihok ingon usa ka kusgan nga ahente nga anti-inflammatory apan usab ang mga metabolite niini.
Ang makapahubag nga mga tigpataliwala ug mga enzyme nga gipugngan sa Nrf2
Mga Cytokine ug Chemokine
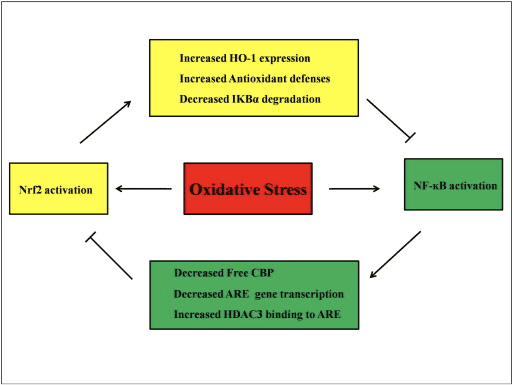
Ang mga cytokine kay ubos ang gibug-aton sa molekula nga mga protina ug mga polypeptide nga gitago sa lainlaing mga selula; sila nag-regulate sa pagtubo sa selula, pagkalahi, ug immune function, ug nalangkit sa panghubag ug pag-ayo sa samad. Ang mga cytokine naglakip sa interleukins (ILs), interferon, tumor necrosis factor (TNF), colony-stimulating factor, chemokines, ug growth factor. Ang ubang mga cytokine giisip nga pro-inflammatory mediators samtang ang uban adunay anti-inflammatory functions. Ang pagkaladlad sa oxidative stress moresulta sa sobrang produksiyon sa mga cytokine nga maoy hinungdan sa oxidative stress sa target cells. Ubay-ubay nga mga pro-inflammatory cytokine ang sobra nga naprodyus kung ang NF-?B gi-activate sa oxidative stress. Dugang pa, ang pro-inflammatory oxidative stress hinungdan sa dugang nga pagpaaktibo sa NF-?B ug ang sobrang produksyon sa mga cytokine. Ang pagpaaktibo sa sistema sa Nrf2 / ARE adunay hinungdanon nga papel sa pagsamok niini nga siklo. Ang mga chemokines usa ka pamilya sa gagmay nga mga cytokine, ang panguna nga papel niini mao ang paggiya sa paglalin sa mga selula sa panghubag. Nag-una kini nga mga chemoattractant alang sa mga leukocytes, monocytes, neutrophils, ug uban pang mga effector cells.
Gikataho nga ang pagpaaktibo sa Nrf2 nagpugong sa LPS-induced transcriptional upregulation sa pro-inflammatory cytokines, lakip ang IL-6 ug IL-1? [50]. IL-1? ug ang produksiyon sa IL-6 nadugangan usab sa Nrf2?/? mga ilaga nga adunay dextran sulfate-induced colitis [51], [52]. Gipugngan sa Nrf2 ang paghimo sa downstream IL-17 ug uban pang mga makapahubag nga mga hinungdan nga Th1 ug Th17, ug gipugngan ang proseso sa sakit sa usa ka eksperimento nga modelo sa multiple sclerosis, autoimmune encephalitis [53]. Ang Nrf2-dependent anti-oxidant genes HO-1, NQO-1, Gclc, ug Gclm block TNF-?, IL-6, monocyte chemo attractant protein-1 (MCP1), macrophage inflammatory protein-2 (MIP2), ug makapahubag mga tigpataliwala. Apan sa kaso sa Nrf2-knockout nga mga ilaga, ang anti-inflammatory nga epekto dili mahitabo [54]. Ang peritoneal neutrophils gikan sa Nrf2-knockout nga mga ilaga nga gitambalan sa LPS adunay mas taas nga lebel sa cytokines (TNF-? ug IL-6) ug chemokines (MCP1 ug MIP2) kay sa wild-type (WT) nga mga selula [54]. Sa vitro, ang pagbalhin sa Nrf2 gene ngadto sa tawo ug rabbit aortic smooth muscle cells makapugong sa pagtago sa MCP1 [8], [55], ug ang Nrf2-dependent nga HO-1 nga ekspresyon nagpugong sa TNF-?-stimulated NF-?B ug MCP-1 pagtago sa umbilical vein endothelial cells sa tawo [56]. Kini nga mga nahibal-an nagpakita nga, isip tubag sa makapahubag nga stimuli, ang upregulation sa Nrf2 signaling makapugong sa sobrang produksyon sa mga pro-inflammatory cytokines ug chemokines ingon man usab sa paglimit sa pagpaaktibo sa NF-?B.
Mga Molecule sa Adhesion sa Cell
Ang cell adhesion molecules (CAMs) maoy mga protina nga nagbugkos sa mga selula o sa extracellular matrix. Nahimutang sa ibabaw sa selula, sila nalangkit sa pag-ila sa selula, pagpaaktibo sa selula, pagbalhin sa signal, pagdaghan, ug pagkalahi. Lakip sa mga CAM, ang ICAM-1 ug VCAM-1 mga importanteng miyembro sa immunoglobulin superfamily. Ang ICAM-1 anaa sa ubos nga konsentrasyon sa leukocyte ug endothelial cell lamad. Sa stimulation sa cytokine, ang konsentrasyon sa kamahinungdanon nagdugang. Ang ICAM-1 mahimong maaghat sa IL-1 ug TNF ug gipahayag sa vascular endothelium, macrophage, ug lymphocytes. Kini usa ka ligand alang sa integrin, usa ka receptor nga makita sa mga leukocyte. Sa diha nga ang ICAM-1-integrin nga tulay gi-activate, ang mga leukocyte mogapos sa mga endothelial cells ug dayon molalin ngadto sa subendothelial tissues [57]. Ang VCAM-1 nagpataliwala sa pagdikit sa mga lymphocytes, monocytes, eosinophils, ug basophils ngadto sa vascular endothelium ug nakatampo sa pagrekrut sa leukocyte, nga sa katapusan mosangpot sa kadaot sa tisyu tungod sa oxidative stress. Gipugngan sa Nrf2 ang kalihokan sa promosyon sa VCAM-1 [58]. Ang Nrf2-regulated downstream gene HO-1 mahimong makaapekto sa ekspresyon sa E-selectin ug VCAM-1, adhesion molecules nga nalangkit sa endothelial cells [59]. Ang pulmonary nga ekspresyon sa daghang mga CAM sama sa CD-14, TREM1, SELE, SELP, ug VCAM-1 mas taas sa Nrf2?/? mga ilaga kay sa Nrf2+/+ nga mga ilaga [60]. Ang Nrf2 sa human aortic endothelial cells nagpugong sa TNF-?-induced VCAM-1 nga ekspresyon ug makabalda sa TNF-?-induced monocytic U937 cell adhesion [8]. Ang overexpression sa Nrf2 usab nagpugong sa TNF-?-induced VCAM-1 gene expression sa human microvascular endothelial cells [61]. Ang natural nga antioxidant nga 3-hydroxyanthranilic acid (HA), usa sa l-tryptophan metabolites nga naporma sa vivo ubay sa metaboliko nga ruta nga nailhan nga kynurenine nga agianan sa panahon sa panghubag o impeksyon, nakit-an nga nagpukaw sa HO-1 nga ekspresyon ug aron mapukaw ang Nrf2 sa umbilical sa tawo. vein endothelial cells (HUVECs). Ang Nrf2-dependent nga HO-1 nga ekspresyon nga gipahinabo sa HA nagpugong sa pagtago sa MCP-1, ekspresyon sa VCAM-1 ug pagpaaktibo sa NF-kB nga may kalabutan sa kadaot sa vascular ug panghubag sa atherosclerosis [56]. Ang anti-proliferative ug anti-inflammatory synthetic chalcone derivative 2?,4?,6?-tris (methoxymethoxy) chalcone nagpugong sa ICAM-1, ang pro-inflammatory cytokine IL-1?, ug TNF-? ekspresyon sa colonic tissue gikan sa mga ilaga nga gitambalan sa trinitrobenzene sulfonic acid [62]. Ang pag-uswag sa Nrf2 nagpugong sa TNF-?-induced ICAM-1 nga ekspresyon sa human retinal pigment epithelial cells nga gitambalan sa lycopene [63]. Ang tanan nga kini nga mga pagtuon nagsugyot nga ang Nrf2 adunay hinungdanon nga papel sa proseso sa panghubag pinaagi sa pag-regulate sa paglalin ug paglusot sa mga selula sa panghubag sa naghubag nga tisyu.
Matrix Metalloproteinases (MMPs)
Ang mga MMP kaylap nga anaa sa extracellular matrix ug nalambigit sa mga proseso sa physiological ug pathological sama sa pagdaghan sa selula, paglalin, pagkalahi, pag-ayo sa samad, angiogenesis, apoptosis, ug metastasis sa tumor. Gikataho nga ang Nrf2 / HO-1 axis nagpugong sa MMP-9 sa mga macrophage ug MMP-7 sa mga selula sa epithelial sa tinai sa tawo, ug kini mapuslanon sa pagtambal sa sakit sa panghubag sa panghubag [62], [64]. Ang kadaot sa panit nga gipahinabo sa UV irradiation mas grabe sa Nrf2-knockout kay sa WT nga mga ilaga ug ang lebel sa MMP-9 mas taas, nga nagpakita nga ang Nrf2 nagpamenos sa MMP-9 nga ekspresyon. Busa, ang Nrf2 giisip nga panalipod batok sa UV irradiation [65]. Ang laing pagtuon usab nagtaho nga ang downregulated transcriptional activation sa MMP-9 sa tumor cell invasion ug panghubag gi-regulate pinaagi sa pagpugong sa NF-kB signaling pathway [66]. Sa traumatic spinal cord injury, ang NF-kB signaling pathway nag-apil usab sa pag-regulate sa lebel sa mRNA sa MMP-9 [67]. Busa, sa panghubag ang regulasyon sa mga MMP direkta nga apektado sa Nrf2 nga agianan o dili direkta pinaagi sa Nrf2-naimpluwensyahan nga NF-?B nga agianan.
Cyclooxygenase-2 (COX2) ug Inducible Nitric Oxide Synthase (INOS)
Ang usa ka serye sa mga eksperimento sa Nrf2-knockout nga mga ilaga nagpakita sa mahinungdanong papel niini sa panghubag ug ang regulasyon sa pro-inflammatory nga mga gene sama sa COX-2 ug iNOS. Sa unang higayon, si Khor et al. nagtaho sa dugang nga ekspresyon sa pro-inflammatory cytokines sama sa COX-2 ug iNOS sa colonic tissues sa Nrf2?/? mga ilaga kon itandi sa WT Nrf2 +/+ nga mga ilaga, nga nagpakita nga ang Nrf2 nagpugong sa ilang kalihokan [51]. Ang laing taho sa pretreatment nga adunay sulforaphane, usa sa iladong Nrf2 activators nga anaa sa cruciferous nga mga utanon, nagpakita sa iyang anti-inflammatory nga epekto sa pagpugong sa ekspresyon sa TNF-?, IL-1?, COX-2, ug iNOS sa mRNA. ug lebel sa protina sa nag-unang peritoneal macrophage gikan sa Nrf2+/+ nga mga ilaga kon itandi niadtong gikan sa Nrf2?/? mga ilaga [68]. Sa susama, ang hippocampus sa Nrf2-knockout nga mga ilaga nga adunay LPS-induced nga panghubag nagpakita usab sa mas taas nga pagpahayag sa mga marker sa panghubag sama sa iNOS, IL-6, ug TNF-? kay sa WT nga mga ilaga [69]. Ingon usab, ang Nrf2-knockout nga mga ilaga sobra ka sensitibo sa oxidative stress nga gipahinabo sa 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine ingon man nagpakita sa dugang nga lebel sa mRNA ug protina sa mga marker sa panghubag sama sa COX-2, iNOS , IL-6, ug TNF-? [70]. Dugang pa, ang mga atay gikan sa Nrf2?/? Ang mga ilaga nga gihagit sa usa ka methionine- ug choline-deficient nga pagkaon adunay ~ 5-fold nga mas taas nga mRNA nga ekspresyon sa Cox2, ug iNOS kay sa gikan sa WT nga mga ilaga sa samang pagkaon, nga nagsugyot sa usa ka anti-inflammatory nga papel sa Nrf2 [71]. Bag-ohay lang, si Kim et al. nagpakita nga ang phytochemical ethyl pyruvate naggamit sa iyang anti-inflammatory ug anti-oxidative nga mga epekto pinaagi sa pagkunhod sa ekspresyon sa iNOS pinaagi sa Nrf2 signaling sa BV2 cells. Gipakita nila nga ang ethyl pyruvate nag-aghat sa nukleyar nga translocation sa Nrf2, nga sa katapusan nagpugong sa interaksyon tali sa p65 ug p300, nga nagdala sa pagkunhod sa ekspresyon sa iNOS [72]. Dugang pa, ang carbazole analogue LCY-2-CHO nagpalihok sa Nrf2 ug nagpahinabo sa nukleyar nga translokasyon niini, nga misangpot sa pagsumpo sa COX2 ug iNOS nga ekspresyon [73] sa rat aortic vascular smooth muscle cells.
Paradoxical nga Papel sa Nrf2 sa Regulasyon sa NLRP3 iIflammasome�Kalihokan
Ang NLR family, pyrin domain nga adunay 3 (NLRP3) inflammasome usa ka multiprotein complex nga naglihok isip pathogen recognition receptor (PRR) ug nag-ila sa halapad nga microbial, oxidative stress signal sama sa pathogen-associated molecular patterns (PAMPs), Damage- kaubang molekular nga mga molekula sa pattern (DAMPs) ug ROS [74]. Ang gi-activate nga NLRP3 inflammasome nagpataliwala sa cleavage sa caspase-1 ug pagtago sa pro-inflammatory cytokine interleukin-1? (IL-1?) nga sa katapusan nag-aghat sa proseso sa pagkamatay sa selula nga nailhan nga pyroptosis nga nanalipod sa mga host batok sa daghang mga pathogens [75]. Bisan pa, ang aberrant nga pagpaaktibo sa inflammasome nalangkit sa mga sakit nga dili maayo sa protina sama sa transmissible spongiform encephalopathies, Alzheimer's disease, Parkinson's disease ug type 2 diabetes [76], cancer [77], gout, ug atherosclerosis [78].
Ang usa ka bag-o nga obserbasyon gikan sa Rong Hu nga grupo sa asosasyon sa Nrf2 nga adunay negatibo nga regulasyon sa inflammasome nagpadayag nga, ang Nrf2 nag-aghat sa NQO1 nga ekspresyon nga mosangpot sa pagdili sa NLRP3 inflammasome activation, caspase-1 cleavage ug IL-1? henerasyon sa mga macrophage. Dugang pa, usa ka ilado nga Nrf2 activator, tert-butylhydroquinone (tBHQ) negatibo nga gikontrol ang NLRP3 transcription pinaagi sa pagpaaktibo sa ARE pinaagi sa Nrf2-dependent nga paagi [79]. Dugang pa sa obserbasyon sa ibabaw, ang sama nga grupo gipadayag usab nga, ang dimethyl fumarate (DMF) nagpugong sa DSS-induced colitis pinaagi sa pagpaaktibo sa Nrf2 signaling pathway nga nalangkit sa Nrf2 nuclear translocation ug inhibition sa NLRP3 inflammasome assembly [80].
Usa ka serye sa mga eksperimento gamit ang natural ug sintetikong mga compound nagpadayag usab sa pagpugong nga epekto sa Nrf2 sa NLRP3 inflammasome activation. Pananglitan, ang pagtambal sa epigallocatechin-3-gallate (EGCG) sa lupus nephritis nga mga ilaga nagpakita sa pagkunhod sa renal NLRP3 inflammasome activation nga gipataliwala sa Nrf2 signaling pathway [81]. Sa samang paagi, ang citral (3,7-dimethyl-2,6-octadienal), usa ka mayor nga aktibong compound sa Chinese herbal medicine nga Litsea cubeba, nagpugong sa NLRP3 inflammasome activation pinaagi sa Nrf2 antioxidant signaling pathway sa Accelerated and Severe Lupus Nephritis (ASLN) mouse model. [82]. Sa susama, ang biochanin gipanalipdan batok sa LPS / GalN-induced liver injury pinaagi sa pagpaaktibo sa Nrf2 pathway ug pagpugong sa NLRP3 inflammasome activation sa laki nga BALB / c mice [83]. Dugang pa, ang mangiferin gipakita usab sa pag-regulate sa ekspresyon sa Nrf2 ug HO-1 sa usa ka paagi nga nagsalig sa dosis ug gipugngan ang LPS / D-GalN-induced hepatic NLRP3, ASC, caspase-1, IL-1? ug TNF-? ekspresyon [84].
Bisan pa sa negatibo nga regulasyon sa NLRP3 sa Nrf2, gipalihok usab niini ang NLRP3 ug AIM2 inflammasome function. Si Haitao Wen ug mga kauban nakadiskobre niana, Nrf2 ?/? Ang mga macrophage sa mouse nagpakita sa depekto nga pagpaaktibo sa NLRP3 ug AIM2 Inflammasome apan dili ang NLRC4 inflammasome [85]. Makaiikag, kini nga obserbasyon naghulagway sa wala mailhi nga mga gimbuhaton sa Nrf2 sa konteksto sa mga sakit nga may kalabutan sa panghubag; busa importante kaayo nga tun-an pa aron ipadayag ang mekanismo diin ang Nrf2 nagpalihok sa inflammasome function sa dili pa kini isipon nga therapeutic target.
Pagpugong sa Pro-Inflammatory Cytokine Transcription ni Nrf2
Usa ka bag-o nga imbestigasyon base sa chromatin immunoprecipitation (ChIP) -seq ug ChIP-qPCR nga mga resulta sa mouse macrophage nagpadayag nga ang Nrf2 nagbugkos sa mga rehiyon sa promoter sa mga pro-inflammatory cytokine sama sa IL-6 ug IL-1? ug nagpugong sa pagrekrut sa RNA Pol II. Ingon nga resulta, ang RNA Pol II dili makahimo sa pagproseso sa transcriptional activation sa IL-6 ug IL-1? nga sa katapusan mosangpot sa pagdili sa ekspresyon sa gene. Sa unang higayon, ang grupo ni Masayuki Yamamoto nagpadayag sa nobela nga mekanismo diin ang Nrf2 dili lamang nag-transactivate sa mga downstream nga mga gene pinaagi sa AREs apan nagpugong usab sa transcriptional nga pagpaaktibo sa mga piho nga mga gene nga adunay o walay ARE pinaagi sa pagpugong sa pagrekrut sa RNA Pol II [50].
Crosstalk Taliwala sa Nrf2 ug NF-?B Pathways
Ang NF-?B usa ka komplikadong protina nga responsable sa transkripsyon sa DNA nga makita sa halos tanang matang sa selula sa mananap ug nalambigit sa nagkalain-laing proseso sama sa panghubag, apoptosis, pagtubag sa imyunidad, pagtubo sa selula, ug paglambo. Ang p65, usa ka Rel protein sa NF-?B nga pamilya, adunay transactivation domain samtang ang p50 wala ug nagkinahanglan og heterodimerization sa Rel protein aron ma-activate ang transcription. Atol sa oxidative stress, ang I?B kinase (IKK) gi-activate ug maoy hinungdan sa phosphorylation sa I?B, nga miresulta sa pagpagawas ug nuclear translocation sa NF-?B. Ang NF-?B maoy hinungdan sa transcription sa mga pro-inflammatory mediator sama sa IL-6, TNF-?, iNOS, IL-1, ug intracellular adhesion COX-2.
Ang abnormal nga regulasyon sa NF-?B konektado sa rheumatoid arthritis, hika, sakit sa panghubag sa tinai, ug Helicobacter pylori infection-induced gastritis [86]. Sa pagkakaron giisip nga ang kalihokan sa NF-kB nag-impluwensya sa Keapl/Nrf2/ARE signaling pathway nag-una sa tulo ka aspeto: una, ang Keap1 nagpaubos sa IKK? pinaagi sa ubiquitination, sa ingon nagpugong sa kalihokan sa NF-?B [87]. Ikaduha, ang proseso sa panghubag nag-aghat sa mga tigpataliwala sa panghubag sama sa COX2 nga nakuha gikan sa cyclopentenone prostaglandin 15d-PGJ2, usa ka lig-on nga electrophile nga nag-reaksyon sa Keap1 ug nagpalihok sa Nrf2, sa ingon nagsugod ang transkripsyon sa gene nga adunay dungan nga pagpugong sa kalihokan sa NF-kB [58], [88] ( Fig. 3 A, B). Ikatulo, ang NF-?B mahimong maghiusa sa kompetisyon nga Nrf2 transcriptional co-activator CBP [89], [90] (Fig. 3 C, D).
Figure 3 Crosstalk tali sa Nrf2 ug NF-?B nga mga agianan. (A) Gimandoan sa Keap1 ang IKK sa CUL3-mediated ubiquitination ug proteasome degradation nga sa katapusan mosangpot sa pagdili sa NF-?B phosphorylation ug kini nga mekanismo naglihok usab isip competitive binding sa Nrf2 ug IKK sa Keap1. (B) Ang stress sa oxidative nagpalihok sa IKK nga nag-phosphorylate sa NF-?B, nga nagpadulong sa paghubad niini ngadto sa nucleus ug pagpaaktibo sa mga proinflammatory cytokine sama sa COX-2. Ang terminal nga produkto sa COX-2 nga nailhan nga 15d-PGJ2 naglihok isip usa ka inducer sa Nrf2 nga sa katapusan mosangpot sa pagsumpo sa oxidative stress. (C) Nrf2 nagbugkos sa iyang transcriptional cofactor CBP uban sa gamay nga Maf ug uban pang transcriptional makinarya sa pagsugod sa ARE-driven gene ekspresyon. (D) Kung ang NF-?B nagbugkos sa CBP sa usa ka kompetisyon nga paagi, kini nagpugong sa pagbugkos sa CBP sa Nrf2, nga mosangpot sa pagdili sa Nrf2 transactivation.
Gituohan nga ang Nrf2 ug NF-?B signaling pathways nakig-interact aron makontrol ang transcription o function sa downstream target proteins. Sa katarungan niini nga pangagpas daghang mga pananglitan nagpakita nga direkta o dili direkta nga pagpaaktibo ug pagdili mahitabo tali sa mga miyembro sa Nrf2 ug NF-?B nga mga agianan (Fig. 4). Agig tubag sa LPS, ang Nrf2 knockdown mahinungdanon nga nagdugang sa NF-?B transcriptional nga kalihokan ug NF-?B-dependent nga transcription sa gene, nga nagpakita nga ang Nrf2 nagpugong sa NF-?B nga kalihokan [60], [91]. Dugang pa, ang dugang nga ekspresyon sa Nrf2-depende sa ubos nga HO-1 nagpugong sa kalihokan sa NF-?B. Kung ang mga selula sa kanser sa prostate maladlad sa makadiyot sa ?-tochopheryl succinate, usa ka gigikanan sa bitamina E, HO-1 nga ekspresyon gi-upregulate. Ang katapusan nga mga produkto sa HO-1 nagpugong sa nukleyar nga translokasyon sa NF-?B [92]. Kini nga mga pagtuon sa vivo nagsugyot nga ang Nrf2 negatibo nga nag-regulate sa NF-kB signaling pathway. LPS stimulates NF-?B DNA binding nga kalihokan ug ang lebel sa p65 subunit sa NF-?B mao ang kamahinungdanon mas taas sa nuclear extracts gikan sa mga baga sa Nrf2?/? kay sa WT nga mga ilaga, nga nagsugyot og negatibo nga papel sa Nrf2 sa NF-?B activation. Dugang pa, Nrf2?/? mouse embryo fibroblasts gitambalan sa LPS ug TNF-? ipakita ang mas prominenteng NF-?B activation tungod sa IKK activation ug I?B-? pagkadaot [60]. Ug ang clearance sa respiratory syncytial virus labi nga mikunhod samtang ang NF-?B DNA-binding nga kalihokan nagdugang sa Nrf2?/? mga ilaga kon itandi sa WT nga mga ilaga [93]. Pristane-induced lupus nephritis sa Nrf2?/? Ang mga ilaga nga gitambalan sa sulforaphane adunay grabe nga kadaot sa bato ug mga pagbag-o sa patolohiya ingon man ang taas nga ekspresyon sa iNOS ug pagpaaktibo sa NF-B kumpara sa WT, nga nagsugyot nga ang Nrf2 nagpauswag sa lupus nephritis pinaagi sa pagpugong sa NF-B nga agianan sa pagsenyas ug paghawan sa ROS [94] ]. Ang kalihokan sa NF-?B mahitabo usab kung ang mga selula gitambalan sa usa ka Nrf2 inducer kauban ang LPS ug TNF-?. Pananglitan, ang usa ka sintetikong chalcone derivative nagpugong sa TNF-?-induced NF-?B nga pagpaaktibo sa direkta ug dili direkta ug sa bahin pinaagi sa induction sa HO-1 nga ekspresyon sa human intestinal epithelial HT-29 cells [62]. Ang pagsumpo sa NF-?B translocation ug DNA-binding nga kalihokan ingon man ang pagsumpo sa iNOS nga ekspresyon sa mga hepatocytes makita kung ang F344 nga mga ilaga gitambalan sa 3H-1,2-dithiole-3-thione (D3T) [95]. Human sa co-treatment sa sulforaphane ug LPS, ang LPS-induced nga ekspresyon sa iNOS, COX-2, ug TNF-? sa Raw 264.7 nga mga macrophage gipaubos, nagsugyot nga ang sulforaphane adunay anti-inflammatory nga kalihokan pinaagi sa pagpugong sa NF-?B DNA binding [96]. Bisan kung gihimo ang daghang mga eksperimento nga mga pagtuon aron ipatin-aw ang kalambigitan tali sa mga agianan sa Nrf2 ug NF-?B, nagpabilin nga nagkasumpaki nga mga resulta. Parehong positibo ug negatibo nga mga regulasyon ang gitaho tali sa Nrf2 ug NF-kB [97]. Kasagaran, ang chemopreventive electrophiles 3H-1,2-dithiole-3-thione, sulforaphane ug Triterpenoid CDDO-Me nagpalihok sa Nrf2 pinaagi sa pagpugong sa NF-kB ug sa mga downregulated nga mga gene niini [98], [99], [100]. Sa kasukwahi, daghang mga ahente o kondisyon sama sa ROS, LPS, flow shear stress, oxidized LDL, ug aso sa sigarilyo gipakita nga nagdugang sa Nrf2 ug NF-kB nga kalihokan [97]. Dugang pa, ang mga pagtuon sa vivo nagpadayag nga ang kalihokan sa NF-kB mikunhod sa mga atay nga nahimulag gikan sa Nrf2?/? Ang mga ilaga ug NF-?B nagbugkos nga kalihokan mas ubos sa Nrf2?/? kay sa Nrf2+/+ nga mga ilaga [101]. Bisan pa, ang mga selula sa aortic endothelial sa tawo nga gitambalan nga adunay adenoviral vector Nrf2 nagpugong sa NF-?B nga mga gene sa ubos nga wala makaapekto sa kalihokan sa NF-?B [8].
Figure 4 Regulatory loop sa Nrf2 ug NF-?B. Ang Nrf2 nga agianan nagpugong sa pagpaaktibo sa NF-?B pinaagi sa pagpugong sa pagkadaot sa I?B-? ug pagdugang sa HO-1 nga ekspresyon ug antioxidant nga mga depensa nga nag-neutralize sa ROS ug detoxifying nga mga kemikal. Ingon nga resulta, ang ROS-associated NF-?B activation gipugngan. Ingon usab, ang NF-?B-mediated transcription nagpamenos sa pagpaaktibo sa Nrf2 pinaagi sa pagkunhod�HUSAY�gene transcription ug libre nga CREB binding protein pinaagi sa pagpakigkompetensya sa Nrf2 para sa CBP. Dugang pa, ang NF-?B nagdugang sa pagrekrut sa histone deacetylase (HDAC3) ngadto sa ARE nga rehiyon ug busa ang Nrf2 transcriptional activation gipugngan.
Ang pagpaaktibo sa Nrf2 signaling pathway adunay dakong papel sa pagpahayag sa mga enzyme ug mga gene nga nalangkit sa detoxification sa mga reactive oxidants pinaagi sa pagpausbaw sa antioxidant nga kapasidad sa mga selula sa lawas sa tawo. Samtang daghang mga pagtuon sa panukiduki ang magamit karon, ang mga mekanismo sa regulasyon sa pagpaaktibo sa Nrf2 dili hingpit nga masabtan. Ang usa ka posible nga papel sa Nrf2 signaling pathway sa pagtambal sa panghubag nakit-an usab. Dr. Alex Jimenez DC, CCST Insight
Papel sa Nrf2 sa Makapahubag nga mga Sakit
Gipakita sa mga pagtuon sa vivo nga ang Nrf2 adunay hinungdanon nga papel sa makapahubag nga mga sakit nga nakaapekto sa lainlaing mga sistema; kini naglakip sa gastritis, colitis, arthritis, pneumonia, kadaot sa atay, sakit sa cardiovascular, sakit nga neurodegenerative, ug kadaot sa utok. Niini nga mga pagtuon, Nrf2?/? mga mananap nagpakita sa mas grabe nga mga sintomas sa panghubag ug tissue kadaot kay sa WT mananap. Busa, gituohan nga ang Nrf2 signaling pathway adunay panalipod nga epekto sa makapahubag nga mga sakit. Ang pag-install sa intra-tracheal sa porcine pancreatic elastase nagpahinabog laygay nga obstructive pulmonary nga sakit, labi na ang emphysema. Ang kulang sa Nrf2 nga mga ilaga dali kaayo sa emphysema, ug ang pagkunhod sa ekspresyon sa HO-1, PrxI, ug ang antiprotease gene SLPI mahitabo sa alveolar macrophage. Ang Nrf2 giisip nga usa ka yawe nga regulator sa macrophage mediated defense system batok sa kadaot sa baga [102]. Ang Nrf2-kulang nga mga ilaga nga adunay emphysema nga gipahinabo sa pagkaladlad sa aso sa tabako sulod sa 6 ka bulan nagpakita sa dugang nga bronchoalveolar nga panghubag, gipataas nga pagpahayag sa oxidative stress marker sa alveoli, ug nadugangan nga alveolar septal cell apoptosis, nga nagsugyot nga ang Nrf2 naglihok batok sa tabako nga gipahinabo sa emphysema pinaagi sa dugang nga ekspresyon sa antioxidant. mga gene [102], [103]. Uban sa Nrf2 disruption, allergen-mediated airway inflammation ug asthma gamit ang ovalbumin complex nagpakita sa dugang nga airway inflammation, airway hyper-reactivity, hyperplasia sa goblet cells, ug taas nga lebel sa Th2 sa bronchoalveolar lavage ug splenocytes, samtang ang Nrf2-mediated signaling pathway naglimite sa airway eosinophilia , mucus hypersecretion, ug airway hyper-reactivity ingon man usab sa pag-aghat sa daghang antioxidant nga mga gene nga makapugong sa pagpalambo sa hika [104]. Ang pag-injection sa carrageenan ngadto sa pleural nga lungag nag-aghat sa pleurisy, ug ang 15d-PGJ2 nga akumulasyon sa Nrf2 nga mga selula sa panghubag kay limitado sa mouse peritoneal macrophage. Sa sayong bahin sa panghubag, ang 15d-PGJ2 nagpalihok sa Nrf2 ug nag-regulate sa proseso sa panghubag pinaagi sa induction sa HO-1 ug PrxI. Gisugyot usab sa usa ka pagtuon nga ang COX-2 adunay anti-inflammatory nga epekto sa sayong bahin pinaagi sa paghimo sa 15d-PGJ2 [105]. Ang oral nga administrasyon sa 1% dextran sulfate sodium sulod sa 1 ka semana nag-aghat sa colitis nga may kalabutan sa histological nga mga pagbag-o nga naglakip sa pagpamubo sa mga crypts ug paglusot sa makapahubag nga mga selula sa colon tissue. Aron mapanalipdan ang intestinal integrity sa colitis, ang Nrf2 mahimong adunay importante nga papel pinaagi sa pag-regulate sa mga pro-inflammatory cytokines ug pag-induce sa phase II detoxifying enzymes [51]. Sa usa ka Nrf2-knockout mouse nga modelo sa LPS-induced pulmonary sepsis, ang NF-?B nga kalihokan nag-regulate sa impluwensya sa makapahubag nga mga cytokine sama sa COX-2, IL-113, IL-6, ug TNF? nga hinungdanon alang sa pagsugod ug pagpasiugda sa panghubag [60]. Gipamenos sa Nrf2 ang kadaot sa panghubag pinaagi sa pag-regulate niining mga hinungdan sa panghubag. Niini nga mga modelo sa acute nga panghubag, ang dugang nga regulasyon sa antioxidant enzymes, pro-inflammatory cytokines, ug mga tigpataliwala sa Nrf2 signaling pathway makapakunhod sa makapahubag nga kadaot sa mga mananap sa WT. Makapainteres, kini usab gitaho sa Nrf2-knockout nga mga ilaga diin ang mga sintomas labi nga gipasamot kon itandi sa WT nga mga ilaga.
Pagpanukiduki sa Nrf2-Dependent Anti-Inflammatory Drugs
Sa katingbanan, among gihisgutan ang mga eksperimento nga nagpakita nga ang Nrf2 signal pathway adunay usa ka regulatory role sa daghang mga dapit sa panghubag, mao nga ang Nrf2-dependent nga anti-inflammatory agent importante alang sa pagtambal sa mga sakit nga makapahubag.
Ang mga tanum labi ka daghan nga gigikanan sa mga compound nga nag-aktibo sa Nrf2 transcription factor, nga nagdala ngadto sa up-regulation sa cytoprotective genes. Bag-ohay lang, daghang mga pagtuon ang gihimo aron masusi ang mga epekto sa lainlaing mga ahente nga anti-inflammatory, kasagaran gikan sa tanum. Pananglitan, ang curcumin mao ang aktibong sangkap sa turmeric ug makita usab sa gamay nga kantidad sa luya; isothiocyanates, ilabi na ang phenylisothiocyanates gikan sa broccoli, celery, ug uban pang mga utanon; ug anthocyanin gikan sa berries ug ubas [124]. Gipakita sa mga pagtuon nga kining tanan nga mga ahente dili lamang maayo nga mga antioxidant apan adunay usab kusog nga anti-inflammatory effect pinaagi sa Nrf2 induction [125], [126]. Busa, ang pag-uswag sa bag-ong anti-inflammatory Nrf2 activators gikan sa extract sa tanom nakadani ug daghang interes sa medikal nga panukiduki.
Sa bag-ohay nga mga tuig, daghang mga eksperimento sa hayop ang gihimo aron makumpirma ang mga aksyon sa kini nga mga compound. Ang artesunate kay gigamit ilabina alang sa grabeng malaria, cerebral malaria, ug rheumatic autoimmune nga mga sakit; epektibo usab kini sa septic lung injury. Ang Artesunate nagpalihok sa Nrf2 ug HO-1 nga ekspresyon, ug ang naulahi nagpamenos sa pag-agos sa pro-inflammatory cytokines ug leukocytes ngadto sa tisyu aron mapugngan ang panghubag [127]. Ang Isovitexin, nga gikuha gikan sa mga kasko sa Oryza sativa nga bugas, gituohan nga adunay anti-inflammatory ug antioxidant properties; kini adunay usa ka panalipod nga papel batok sa LPS-induced acute lung injury pinaagi sa pagpaaktibo sa Nrf2 / HO-1 nga agianan ug pagpugong sa MAPK ug NF-?B [128]. Ang Fimasartan, usa ka bag-ong sikat nga angiotensin II receptor blocker nga naglihok sa renin-angiotensin system, nagpamenos sa presyon sa dugo; Ang paggamit sa fimasartan sa pagtratar sa mga ilaga nga adunay surgically-induced unilateral ureteral obstruction makapamenos sa oxidative stress, panghubag, ug fibrosis pinaagi sa upregulating Nrf2 ug ang antioxidant nga agianan ug pagpugong sa RAS ug MAPKs [129]. Ang Sappanone kaylap nga giapod-apod sa Southeast Asia, diin gigamit kini isip anti-influenza, anti-allergic, ug neuroprotective nga tambal; kini nagpalihok sa Nrf2 ug nagpugong sa NF-?B ug busa mahimong mapuslanon sa pagtambal sa Nrf2- ug/o NF-?B-related nga mga sakit [130]. Ang bixin nga gikuha gikan sa mga liso sa Bixin orellana gigamit alang sa makatakod ug makapahubag nga mga sakit sa Mexico ug South America; kini nagpamenos sa makapahubag nga mga mediator, alveolar capillary leakage, ug oxidative nga kadaot sa Nrf2-dependent nga paagi aron sa paghupay sa kadaot sa baga nga gipahinabo sa bentilasyon ug pagpasig-uli sa normal nga morphology sa baga [131]. Ang ubang mga compound sa tanum, sama sa epigallocatechin gallate, sulforaphane, resveratrol, lycopene, ug green tea extract adunay mga therapeutic effect sa makapahubag nga mga sakit pinaagi sa Nrf2 signaling pathway [132], [133], [134]. Bag-ohay lang ang laing phytochemical, eriodictyol, nga anaa sa citrus fruit, gitaho nga adunay anti-inflammatory ug antioxidant nga mga epekto sa cisplatin-induced kidney injury ug sepsis-induced acute lung injury pinaagi sa pag-regulate sa Nrf2, inhibiting NF-?B, ug inhibiting the pagpahayag sa mga cytokine sa macrophage [135], [136]. Bisan pa, daghang mga phytochemical ang nagpakita ug dako nga saad alang sa pagpugong ug pagtambal sa lainlaing mga sakit sa tawo, ug ang uban nakasulod na sa yugto sa mga pagsulay sa klinika (Table 2).
Kini nga mga compound sa tanum nag-aktibo sa Nrf2 signaling pathway sa panguna sa porma sa mga electrophilic nga materyales nga nagbag-o sa mga residu sa cysteine sa Keap1, nga nagpadulong sa libre nga nukleyar nga Nrf2 nga nagbugkos sa ARE, nga miresulta sa pagpaaktibo sa transkripsyon sa katugbang nga gene.
Sulforaphane ug ang mga Epekto Niini sa Kanser, Pagka-mortal, Pagkatigulang, Utok ug Panggawi, Sakit sa Kasingkasing ug uban pa
Ang Isothiocyanates mao ang pipila sa labing hinungdanon nga mga compound sa tanum nga makuha nimo sa imong pagkaon. Niini nga video gihimo nako ang labing komprehensibo nga kaso alang kanila nga nahimo sukad. Mubo nga gidugayon sa pagtagad? Laktaw sa imong paborito nga hilisgutan pinaagi sa pag-klik sa usa sa mga punto sa oras sa ubos. Full timeline sa ubos.
Pangunang mga seksyon:
00:01:14 – Kanser ug mortalidad
00:19:04 - Pagkatigulang
00:26:30 - Utok ug pamatasan
00:38:06 - Katapusan nga recap
00:40:27 - Dosis
Tibuok timeline:
00:00:34 - Pagpaila sa sulforaphane, usa ka mayor nga focus sa video.
00:01:14 - Pagkonsumo sa cruciferous nga utanon ug pagkunhod sa tanan nga hinungdan sa pagkamatay.
00:02:12 - Risgo sa kanser sa prostate.
00:02:23 - Risgo sa kanser sa pantog.
00:02:34 - Ang kanser sa baga sa mga nanigarilyo peligro.
00:02:48 - Risgo sa kanser sa suso.
00:03:13 - Hypothetical: unsa man kung ikaw adunay kanser? (interbensyonal)
00:03:35 - Katuohan nga mekanismo nga nagmaneho sa kanser ug mortalidad nga asosasyon nga datos.
00:04:38 - Sulforaphane ug kanser.
00:05:32 - Ang ebidensya sa hayop nga nagpakita sa kusog nga epekto sa broccoli sprout extract sa pag-uswag sa tumor sa pantog sa mga ilaga.
00:06:06 - Epekto sa direktang suplemento sa sulforaphane sa mga pasyente sa kanser sa prostate.
00:07:09 - Bioaccumulation sa isothiocyanate metabolites sa aktwal nga tisyu sa dughan.
00:08:32 - Pagpugong sa mga stem cell sa kanser sa suso.
00:08:53 - Leksyon sa kasaysayan: ang mga brassicas gitukod nga adunay mga kabtangan sa kahimsog bisan sa karaang Roma.
00:09:16 - Ang abilidad sa Sulforaphane sa pagpalambo sa carcinogen excretion (benzene, acrolein).
00:10:10 - Sa unsang paagi ang pagpaaktibo sa NRF2 makapauswag sa carcinogen excretion pinaagi sa glutathione-S-conjugates.
00:10:34 - Ang Brussels sprouts nagdugang sa glutathione-S-transferase ug makapamenos sa kadaot sa DNA.
00:11:20 - Ang broccoli sprout nga ilimnon nagdugang sa benzene excretion sa 61%.
00:13:31 - Ang homogenate nga pagtubo sa broccoli nagdugang antioxidant enzymes sa taas nga agianan sa hangin.
00:15:45 - Pagkonsumo sa cruciferous nga utanon ug pagkamatay sa sakit sa kasingkasing.
00:16:55 - Ang broccoli sprout powder nagpauswag sa mga lipid sa dugo ug sa kinatibuk-ang risgo sa sakit sa kasingkasing sa type 2 nga mga diabetes.
00:19:04 - Pagsugod sa seksyon sa pagkatigulang.
00:19:21 - Sulforaphane-enriched nga pagkaon nagpauswag sa kinabuhi sa mga bakukang gikan sa 15 ngadto sa 30% (sa pipila ka mga kondisyon).
00:20:34 - Ang kamahinungdanon sa ubos nga panghubag alang sa taas nga kinabuhi.
00:22:05 - Ang cruciferous nga mga utanon ug broccoli sprout powder daw makapakunhod sa nagkalainlain nga mga marker sa panghubag sa mga tawo.
00:23:40 - Mid-video recap: kanser, mga seksyon sa pagkatigulang
00:24:14 - Ang mga pagtuon sa mouse nagsugyot nga ang sulforaphane mahimong makapauswag sa adaptive immune function sa pagkatigulang.
00:25:18 - Ang Sulforaphane nagpauswag sa pagtubo sa buhok sa usa ka modelo sa mouse sa pagkalbo. Hulagway sa 00:26:10.
00:26:30 - Pagsugod sa seksyon sa utok ug pamatasan.
00:27:18 - Epekto sa broccoli sprout extract sa autism.
00:27:48 - Epekto sa glucoraphanin sa schizophrenia.
00:28:17 - Pagsugod sa diskusyon sa depresyon (katuohan nga mekanismo ug pagtuon).
00:31:21 - Ang pagtuon sa mouse gamit ang 10 ka lain-laing mga modelo sa stress-induced depression nagpakita sa sulforaphane nga parehas nga epektibo sama sa fluoxetine (prozac).
00:32:00 - Gipakita sa pagtuon nga ang direkta nga pag-inom sa glucoraphanin sa mga ilaga parehas nga epektibo sa pagpugong sa depresyon gikan sa sosyal nga kapildihan nga modelo sa stress.
00:33:01 - Pagsugod sa seksyon sa neurodegeneration.
00:33:30 - Sulforaphane ug Alzheimer's disease.
00:33:44 - Sulforaphane ug Parkinson's disease.
00:33:51 - Sulforaphane ug sakit sa Hungtington.
00:34:13 - Ang Sulforaphane nagdugang sa mga protina sa heat shock.
00:34:43 - Pagsugod sa seksyon sa traumatic brain injury.
00:35:01 - Sulforaphane gi-injected dayon human ang TBI nagpalambo sa memorya (pagtuon sa mouse).
00:35:55 - Sulforaphane ug neuronal plasticity.
00:36:32 - Ang Sulforaphane nagpauswag sa pagkat-on sa modelo sa type II diabetes sa mga ilaga.
00:37:19 - Sulforaphane ug duchenne muscular dystrophy.
00:37:44 - Pagpugong sa Myostatin sa mga selula sa satellite sa kalamnan (in vitro).
00:38:06 - Late-video recap: mortalidad ug kanser, kadaot sa DNA, oxidative stress ug panghubag, benzene excretion, sakit sa cardiovascular, type II diabetes, mga epekto sa utok (depresyon, autism, schizophrenia, neurodegeneration), NRF2 nga agianan.
00:40:27 - Mga hunahuna sa paghunahuna sa usa ka dosis sa broccoli sprouts o sulforaphane.
00:41:01 - Mga anekdota sa pagtubo sa balay.
00:43:14 - Sa temperatura sa pagluto ug kalihokan sa sulforaphane.
00:43:45 - Ang pagkakabig sa bakterya sa gut sa sulforaphane gikan sa glucoraphanin.
00:44:24 - Ang mga suplemento mas maayo kung gihiusa sa aktibo nga myrosinase gikan sa mga utanon.
00:44:56 - Mga teknik sa pagluto ug cruciferous nga mga utanon.
00:46:06 - Isothiocyanates isip goitrogens.
konklusyon
Sa pagkakaron, daghang panukiduki ang naka-focus sa papel sa Nrf2 / Keap1 / ARE signaling pathway sa panghubag. Lakip sa mga enzyme nga gi-upregulate sa Nrf2, ang HO-1 usa sa mga representante sa stress response enzymes. Ang HO-1 adunay prominenteng anti-inflammatory ug antioxidant properties. Sa kinatibuk-an, ang Nrf2 signaling pathway negatibo usab nga nag-regulate sa mga cytokine, chemokine nga nagpagawas sa mga hinungdan, MMPs, ug uban pang makapahubag nga mga mediator nga COX-2 ug iNOS nga produksyon, nga direkta o dili direkta nga makaapekto sa may kalabutan nga NF-kB ug MAPK nga mga agianan ug uban pang mga network nga nagkontrol sa panghubag. Gisugyot nga ang Nrf2 ug NF-?B signaling pathways makig-interact aron ma-regulate ang transcription o function sa downstream target proteins. Ang pagsumpo o dili pagpaaktibo sa NF-?B-mediated transcriptional nga kalihokan pinaagi sa Nrf2 lagmit mahitabo sa sayong bahin sa panghubag, tungod kay ang NF-?B nag-regulate sa de novo synthesis sa usa ka han-ay sa mga pro-inflammatory mediator. Bisan pa, adunay pipila ka mga limitasyon sa panukiduki sama sa kung adunay mga koneksyon tali sa Nrf2 ug uban pang mga agianan sa pagsenyas sama sa JAK / STAT, ang kamahinungdanon sa karon nga mga activator sa Nrf2 nga nakuha gikan sa mga gigikanan sa natural nga tanum sa panghubag, ug kung giunsa ang pagpauswag sa biolohikal nga kalihokan. ug pauswagon ang pagtarget niini nga mga compound. Nagkinahanglan kini og dugang nga eksperimento nga validation.
Dugang pa, ang Nrf2 signaling pathway maka-regulate sa> 600 genes [163], diin> 200 encode cytoprotective proteins [164] nga nalangkit usab sa panghubag, kanser, neurodegenerative nga mga sakit, ug uban pang dagkong mga sakit [165]. Ang nagtubo nga mga ebidensya nga nagsugyot nga, ang Nrf2 signaling pathway gi-deregulated sa daghang mga kanser, nga miresulta sa aberrant nga ekspresyon Nrf2 dependent gene battery. Dugang pa, ang panghubag adunay dakong papel sa mga sakit nga may kalabutan sa oxidative stress labi na sa cancer. Ang paggamit sa ubay-ubay nga Nrf2 activators aron masumpo ang panghubag mahimong moresulta sa aberrant nga ekspresyon sa Nrf2 downstream genes nga nag-aghat sa oncogenesis ug pagbatok sa chemo ug / o radio therapy. Busa, ang labi ka piho nga mga activator sa Nrf2 mahimong maugmad aron mapamenos ang mga epekto sa pleiotropic. Daghang mga activator sa Nrf2 ang nagpakita sa usa ka mahinungdanon nga pag-uswag sa mga anti-inflammatory function sa oxidative stress related nga mga sakit. Ang pinakamaayo nga pananglitan sa Nrf2 activator nga gi-aprobahan sa FDA ug kaylap nga gigamit alang sa pagtambal sa makapahubag nga sakit sama sa Multiple sclerosis (MS) mao ang dimethyl fumarate. Ang Tecfidera (narehistro nga ngalan sa dimethyl fumarate ni Biogen) epektibo nga gigamit sa pagtambal sa nagbalikbalik nga mga porma sa multiple sclerosis sa daghang mga pasyente [152]. Bisan pa, ang kaepektibo sa paggamit sa Nrf2 activators sa pagtambal sa mga sakit nga makapahubag nanginahanglan dugang nga pag-validate aron malikayan ang makadaot nga mga epekto sa Nrf2. Busa, ang pagpalambo sa mga terapiya alang sa anti-inflammation nga kalihokan nga gipataliwad-an sa Nrf2 mahimong adunay mahinungdanon nga klinikal nga epekto. Ang nagpadayon nga mga pagtuon sa Nrf2 signaling pathway sa tibuok kalibutan gigahin sa pagpalambo sa mga gitumong kaayo nga mga ahente sa pagtambal aron makontrol ang mga sintomas sa panghubag, ug aron mapugngan ug matambalan ang kanser ingon man ang neurodegenerative ug uban pang dagkong mga sakit.
Sa konklusyon, ang Nrf2 nakamatikod sa lebel sa oxidative stress sa lawas sa tawo ug sa katapusan makatabang sa pagpalambo sa regulasyon sa antioxidant ug detoxifying enzymes ug genes. Tungod kay ang laygay nga panghubag tungod sa pagtaas sa lebel sa oxidative stress nalambigit sa mga sakit nga neurodegenerative, Ang Nrf2 mahimong adunay hinungdanon nga papel sa pagtambal sa mga isyu sa kahimsog sama sa Alzheimer's disease, ug uban pa. Ang sakup sa among kasayuran limitado sa mga isyu sa kahimsog sa chiropractic ug spinal. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Dugang nga Hisgutan sa Hilisgutan: Paghupay sa Sakit sa Tuhod nga walay Operasyon
Ang sakit sa tuhod usa ka ilado nga simtomas nga mahimong mahitabo tungod sa lain-laing mga samad sa tuhod ug/o kondisyon, lakip nasports injuries. Ang tuhod mao ang usa sa labing komplikado nga mga lutahan sa lawas sa tawo tungod kay kini gilangkoban sa intersection sa upat ka bukog, upat ka ligaments, lain-laing mga tendon, duha ka menisci, ug cartilage. Sumala sa American Academy of Family Physicians, ang kasagarang hinungdan sa kasakit sa tuhod naglakip sa patellar subluxation, patellar tendinitis o jumper's knee, ug Osgood-Schlatter disease. Bisan kung ang sakit sa tuhod lagmit nga mahitabo sa mga tawo nga kapin sa 60 ka tuig ang edad, ang sakit sa tuhod mahimo usab nga mahitabo sa mga bata ug mga tin-edyer. Ang sakit sa tuhod mahimong matambalan sa balay subay sa mga pamaagi sa RICE, bisan pa, ang grabe nga mga samad sa tuhod mahimong magkinahanglan dayon nga medikal nga atensyon, lakip ang pag-atiman sa chiropractic. �
Ang mga sakit nga neurodegenerative, sama sa Alzheimer's disease ug Parkinson's disease, nakaapekto sa milyon-milyong mga indibidwal sa tibuok kalibutan. Ang lainlaing mga kapilian sa pagtambal magamit aron matambal ang mga simtomas sa daghang mga sakit sa neurodegenerative bisan kung ang mga sangputanan kanunay nga limitado. Nakaplagan sa mga pagtuon sa panukiduki nga ang oxidative stress nga gipahinabo sa internal ug external nga mga hinungdan mahimong hinungdan sa pag-uswag sa mga sakit nga neurodegenerative. Ang transcription factor, Nrf2, determinado nga molihok isip usa ka mayor nga mekanismo sa depensa batok sa oxidative stress. Ang katuyoan sa artikulo sa ubos mao ang pagpakita sa mga epekto sa Nrf2 sa mga sakit nga neurodegenerative.
Modulasyon sa Proteostasis pinaagi sa Transcription Factor NRF2
Ang mga sakit nga neurodegenerative nalambigit sa pagtipon sa piho nga mga aggregate sa protina, nga nagsugyot sa usa ka suod nga koneksyon tali sa nasamdan nga utok ug pagkawala sa proteostasis. Ang Proteostasis nagtumong sa tanan nga mga proseso diin ang mga selyula nagkontrol sa kadagaya ug pagpilo sa proteome salamat sa usa ka halapad nga network nga naghiusa sa regulasyon sa mga agianan sa pagsenyas, ekspresyon sa gene ug mga sistema sa pagkadaot sa protina. Kini nga pagrepaso misulay sa pag-summarize sa labing may kalabutan nga mga nahibal-an mahitungod sa transcriptional modulation sa proteostasis nga gihimo sa transcription factor NRF2 (nuclear factor (erythroid-derived 2) -like 2). Ang NRF2 giisip nga klasikal nga master regulator sa pagtubag sa antioxidant cell, bisan kung kini karon mitumaw ingon usa ka hinungdanon nga sangkap sa makinarya sa transduction aron mapadayon ang proteostasis. Sama sa atong hisgutan, ang NRF2 mahimong lantawon isip usa ka hub nga nag-compile sa mga emergency signal nga nakuha gikan sa misfolded protein accumulation aron makatukod og usa ka coordinated ug perdurable transcriptional response. Kini makab-ot pinaagi sa mga gimbuhaton sa NRF2 nga may kalabutan sa pagkontrolar sa mga gene nga nalangkit sa pagmintinar sa endoplasmic reticulum physiology, ang proteasome ug autophagy.
keywords:Neurodegenerative nga mga sakit, Unfolded protein response, Proteasome, Ubiquitin, Autophagy, Oxidative stress
Ang Nuclear Factor (erythroid-derived 2)-like 2 (NRF2) kay usa ka basic-leucine-zipper nga protina nga gikonsiderar karon isip master regulator sa cellular homeostasis. Gikontrol niini ang basal ug stress-inducible nga ekspresyon sa sobra sa 250 ka mga gene nga adunay managsama nga cis-acting enhancer nga gitawag nga antioxidant response element (ARE) [1], [2], [3], [4], [5]. Kini nga mga gene moapil sa phase I, II ug III detoxification reactions, glutathione ug peroxiredoxin/thioredoxin metabolism, NADPH production pinaagi sa pentose phosphate pathway ug malic enzyme, fatty acid oxidation, iron metabolism, ug proteostasis [6]. Tungod niining lapad nga cytoprotective functions, posible nga ang usa ka pharmacological hit sa NRF2 mahimong makapakunhod sa epekto sa mga nag-unang hinungdan sa mga sakit nga malala, lakip ang oxidative, inflammatory ug proteotoxic stress. Ang papel sa NRF2 sa modulasyon sa antioxidant defense ug resolusyon sa panghubag natubag sa daghang mga pagtuon (gisusi sa [7]). Dinhi, atong ipunting ang papel niini sa proteostasis, ie, ang homeostatic control sa synthesis sa protina, pagpilo, trafficking ug degradation. Ang mga pananglitan igahatag sa konteksto sa mga sakit nga neurodegenerative.
Pagkawala sa Proteostasis Impluwensya NRF2 Kalihokan sa Neurodegenerative Sakit
Usa ka kinatibuk-ang timaan sa mga sakit nga neurodegenerative mao ang pagkahitabo sa aberrant aggregation sa pipila nga mga protina. Busa, ang sayop nga pagkapilo sa protina aggregates sa ?-synuclein (?-SYN) makita sa Parkinson's disease (PD), ?-amyloid (A?) plaques ug hyper-phosphorylated TAU neurofibrillary tangles sa Alzheimer's disease (AD), huntingtin (Htt) sa Huntington's disease (HD), superoxide dismutase 1 (SOD1) ug TAR DNA binding protein 43 (TDP-43) sa amyotrophic lateral sclerosis (ALS), prion protein (PrP) sa spongiform encephalopathies, ug uban pa. Ang mga aggregate sa protina mahimong adunay epekto sa pipila mga agianan sa cellular, nga mahimo’g makaapekto sa lebel ug kalihokan sa NRF2.
Lahi nga Layers sa Regulasyon Hugot nga Gikontrol ang Kalihokan sa NRF2
Ubos sa mga kondisyon sa pisyolohikal, ang mga selula nagpakita sa ubos nga lebel sa protina sa NRF2 tungod sa paspas nga pagbalhin niini. Agig tubag sa lainlaing stimuli, ang NRF2 nga protina natipon, mosulod sa nucleus ug nagdugang sa transkripsyon sa mga gene nga adunay ARE. Busa, ang pagdumala sa lebel sa protina sa NRF2 usa ka mahinungdanong punto nga kinahanglan nga maghiusa sa positibo ug negatibo nga mga signal sa input. Samtang atong hisgutan ang dugang, ang NRF2 gi-activate pinaagi sa nagkalain-laing mga overlapping nga mga mekanismo aron sa pag-orchestrate sa usa ka paspas ug episyente nga tubag apan sa laing bahin ang NRF2 mahimong mapugngan, tingali sa ikaduhang hugna, aron sa pagpalong sa iyang tubag.
Gikan sa klasiko nga punto sa panglantaw, ang pagpaaktibo sa NRF2 giisip nga usa ka sangputanan sa cellular nga tubag sa oxidant o electrophilic compounds. Niini nga bahin, ang ubiquitin E3 ligase adapter Kelch-sama sa ECH-associated protein 1 (KEAP1) adunay hinungdanon nga papel. Ang mga detalye sa molekula pagahisgotan pa sa Seksyon 4.1. Sa laktud, ang KEAP1 naglihok isip redox sensor tungod sa kritikal nga cysteine residues nga mosangpot sa NRF2 ubiquitination ug proteasomal degradation. Dugang pa sa kini nga klasiko nga modulasyon, ang NRF2 labi nga gikontrol sa mga panghitabo sa pagsenyas. Sa tinuud, ang lainlaing mga kinase gipakita sa phosphorylate ug pag-regulate sa NRF2. Pananglitan, ang NRF2 mahimong phosphorylated sa mitogen activated protein kinases (MAPKs), bisan kung ang kontribusyon niini sa kalihokan sa NRF2 nagpabilin nga dili klaro [8], [9], [10], [11]. PKA kinase ingon man ang pipila ka PKC isozymes [12], CK2 [13] o Fyn [14] phosphorylate NRF2 nga nagbag-o sa kalig-on niini. Ang miaging trabaho gikan sa among grupo nagtaho nga ang glycogen synthase kinse-3? (GSK-3?) Nagpugong sa NRF2 pinaagi sa nuclear exclusion ug proteasomal degradation [15], [25], [26], [27], [28], [29], [30]. Ang mga detalye sa molekula hisgotan sa Seksyon 4.1. Dugang pa, ang NRF2 gisumite sa ubang mga matang sa regulasyon. Pananglitan, ang NRF2 acetylation sa CBP / p300 nagdugang sa kalihokan niini [17], samtang kini gipugngan sa miR153, miR27a, miR142-5p, ug miR144 [16], o pinaagi sa methylation sa cytosine-guanine (CG) nga mga isla sulod sa NRF2 promoter [18].
Epekto sa Protein Aggregates sa NRF2 Regulatory Mechanisms
Niini nga seksyon atong ipunting kung giunsa ang pagtipon sa sayup nga protina mahimo’g makaapekto sa kalihokan sa NRF2 nga naghatag pipila nga mga agianan nga gihisgutan sa ibabaw ingon mga pananglitan nga ilustrasyon. Una, kinahanglan natong tagdon nga ang pagtipon sa protina hugot nga nalambigit sa kadaot sa oxidative. Sa tinuud, ang sayup nga pagtipon sa protina ug pagtipon nag-aghat sa dili normal nga produksiyon sa reactive oxygen species (ROS) gikan sa mitochondria ug uban pang mga gigikanan [19]. Sama sa gihisgutan sa ibabaw, ang ROS mag-usab sa redox-sensitive cysteinees sa KEAP1 nga mosangpot sa pagpagawas, pag-stabilize ug nukleyar nga lokalisasyon sa NRF2.
Mahitungod sa proteinopathies, usa ka pananglitan sa dysregulated signaling nga mga panghitabo nga mahimong makaapekto sa NRF2 gihatag sa hyperactivation sa GSK-3? sa AD. Ang GSK-3?, nailhan usab nga TAU kinase, miapil sa phosphorylation niining microtubule-associated protein, nga miresulta sa paghugpong niini, pagporma sa neurofibrillary tangles ug pagkabalda sa axonal transport (gisusi sa [20]). Sa laing bahin, GSK-3? mahinuklugong makapamenos sa lebel ug kalihokan sa NRF2 sama sa gihisgotan sa ibabaw. Bisan tuod dili kaylap nga gidawat, ang amyloid cascade nagsugyot nga ang makahilo nga A? Ang mga oligomer nagdugang sa GSK-3? kalihokan kauban ang TAU hyper-phosphorylation ug pagkamatay sa neuron [21], [22]. Adunay lain-laing mga modelo sa pagpatin-aw sa unsa nga paagi A? pabor sa GSK3-? kalihokan. Pananglitan, A? nagbugkos sa insulin receptor ug nagpugong sa PI3K ug AKT signaling pathways, nga hinungdanon sa pagpadayon sa GSK-3? dili aktibo sa phosphorylation sa N-terminal Ser9 residue [23]. Sa laing bahin, extracellular A? nakig-uban sa Frizzled receptors, nagbabag sa WNT signaling [24] ug miresulta pag-usab sa pagpagawas sa aktibo nga GSK-3?. Sa katingbanan, A? akumulasyon modala ngadto sa abnormal hyperactivation sa GSK-3?, sa ingon makadaot sa usa ka tukma nga NRF2 tubag.
Sama sa gihisgutan sa mosunod nga seksyon, ang sayop nga mga protina mosangpot sa pagpaaktibo sa PERK ug MAPKs, nga sa baylo nag-regulate sa NRF2 [31], [8], [9], [10], [11]. Dugang pa, ang dysregulated nga CBP / p300 nga kalihokan gitaho sa daghang mga proteinopathies [32] ug ang usa ka global nga pagkunhod sa DNA methylation sa mga utok sa AD gipakita usab [33], busa naghatag mga sukaranan aron masusi ang kalabotan sa kini nga mga nahibal-an sa regulasyon sa NRF2.
Kami ug ang uban nakakita sa mga necropsies sa mga pasyente sa PD ug AD nga pagtaas sa lebel sa protina sa NRF2 ug pipila sa mga target niini, sama sa heme oxygenase 1 (HMOX1), NADPH quinone oxidase 1 (NQO1), p62, ug uban pa, pinaagi sa immunoblot ug pinaagi sa immunohistochemistry [34], [35], [36], [37], [38], [39]. Ang up-regulasyon sa NRF2 sa niini nga mga sakit gihubad nga usa ka dili malampuson nga pagsulay sa masakiton nga utok aron mabawi ang mga kantidad sa homeostatic. Bisan pa, ang laing pagtuon nagpakita nga ang NRF2 kasagarang na-localize sa cytoplasm sa AD hippocampal neurons, nga nagsugyot sa pagkunhod sa NRF2 transcriptional nga kalihokan sa utok [40]. Mahunahunaan nga ang kalainan sa kini nga mga obserbasyon adunay kalabotan sa mga pagbag-o sa mga hinungdan nga nagkontrol sa NRF2 ubay sa mga progresibong yugto sa neurodegeneration.
Tulo ka dagkong sistema ang nakatampo sa proteostasis, nga mao ang unfolded protein response (UPR), ang ubiquitin proteasome system (UPS) ug autophagy. Sunod, gipresentar namon ang ebidensya aron mahanduraw ang NRF2 ingon usa ka hub nga nagkonektar sa mga signal sa emerhensya nga gi-aktibo sa mga aggregate sa protina nga adunay makina nga gigikanan sa protina.
NRF2 Nag-apil sa Unfolded Protein Response (UPR)
Pag-aktibo sa NRF2 isip Tubag sa UPR
Ang pagpilo sa oxidative nga protina sa ER gimaneho sa daghang lahi nga mga agianan, ang labing gitipigan nga naglakip sa protina disulfide-isomerase (PDI) ug ang sulfhydryl oxidase endoplasmic oxidoreductin 1 (ERO1? ug ERO1? sa mga mammal) ingon disulfide donor. Sa laktod, ang PDI nag-catalyze sa pagporma ug pagkaguba sa disulfide bonds tali sa mga residu sa cysteine sulod sa mga protina, samtang kini napilo, tungod sa pagkunhod ug oksihenasyon sa kaugalingon nga cysteine aminoacids. Ang PDI gi-recycle pinaagi sa aksyon sa housekeeping enzyme ERO1, nga nag-reintroduces sa disulfide bonds ngadto sa PDI [41]. Ang molekular nga oxygen mao ang terminal nga electron acceptor sa ERO1, nga nagpatunghag stoichiometric nga kantidad sa hydrogen peroxide alang sa matag disulfide bond nga gihimo [42]. Ang mga peroxidases (PRX4) ug glutathione peroxidases (GPX7 ug GPX8) maoy mga importanteng enzyme aron makunhuran ang hydrogen peroxide sa ER. Kung kini nga oxido-reductive nga sistema dili molihok sa husto, ang abnormal nga akumulasyon sa misfolded nga mga protina mahitabo sa ER ug usa ka hugpong sa mga signal nga ginganlan nga unfolded protein response (UPR) gipasa ngadto sa cytoplasm ug nucleus aron matukod pag-usab ang ER homeostasis [43]. Tulo ka mga protina nga may kalabotan sa lamad ang nahibal-an alang sa pag-ila sa stress sa ER sa mga eukaryotes: pagpaaktibo sa transcription factor 6 (ATF6), pancreatic ER eIF2? kinase (PERK, usab double-stranded RNA-activated protein kinase-sama sa ER kinase), ug inositol-nga nagkinahanglan kinase1 (IRE1). Ang luminal domain sa matag sensor gigapos sa usa ka 78 kDa chaperone nga gitawag ug glucose-regulated protein (GRP78/BIP). Ang BIP nag-dissociate sa tensiyon sa ER aron mabugkos ang mga protina nga gibuklad, nga misangpot sa pagpaaktibo sa tulo ka mga sensor [44].
Ang NRF2 ug ang homologue niini nga NRF1, nga may kalabutan usab sa pagtubag sa antioxidant, miapil sa pagbalhin sa UPR ngadto sa nucleus. Sa kaso sa NRF1, kini nga protina nahimutang sa ER membrane ug moagi sa nuclear translocation sa deglycosylation o cleavage. Dayon, ang pagpaaktibo sa UPR mosangpot sa pagproseso sa NRF1 ug nukleyar nga pagtipon sa resulta nga tipik sa nukleyar nga kompartamento. Bisan pa, ang abilidad sa pag-transactivate sa mga gene nga adunay ARE niini nga tipik sa NRF1 anaa pa sa diskusyon [45].
Gipakita ni Glover-Cutter ug mga kauban sa trabaho ang pagpaaktibo sa NRF2 orthologue sa C. elegans, SKN-1, nga adunay lainlaing mga stressor sa ER. Ang dugang nga ekspresyon sa SKN-1 nagsalig sa lainlaing mga tigpataliwala sa UPR, lakip ang IRE1 o PERK worm orthologues [46]. Sa mga selula nga kulang sa PERK, ang pagkadaot sa synthesis sa protina mosangpot sa pagtipon sa endogenous peroxide ug sa sunod nga apoptosis [47]. Ang effector nga gigamit sa PERK aron mapanalipdan ang ER gikan niini nga mga peroxide mahimong NRF2, tungod kay gitaho nga ang PERK phosphorylates NRF2 sa Ser40, sa ingon mapugngan ang pagkadaut niini sa KEAP1 [31]. Ang induction sa ASK1 lagmit usab nga adunay papel sa kini nga ruta pinaagi sa TRAF2-mediated kinase nga aksyon sa IRE1 [48]. Bisan tuod ang papel sa MAPKs sa regulasyon sa NRF2 kontrobersyal gihapon, bag-o lang gisugyot nga ang IRE1-TRAF2-ASK1-JNK nga agianan mahimong mag-activate sa NRF2 [49] (Fig. 1). Makaiikag, sa C. elegans ug mga selula sa tawo, ang bag-ong ebidensya nagsugyot nga ang cysteine sulfenylation sa IRE1 kinase sa iyang activation loop nagpugong sa IRE1-mediated UPR ug nagpasiugda sa p38 antioxidant nga tubag nga gimaneho sa NRF2. Gisugyot sa datos nga ang IRE1 adunay karaan nga function ingon usa ka cytoplasmic sentinel nga nagpalihok sa p38 ug NRF2 [50].
Figure 1 Regulasyon sa NRF2 sa UPR. Ang pagtipon sa wala mapilo o sayop nga pagkapilo nga mga protina sulod sa endoplasmic reticulum mahimong makasugod sa unfolded protein response (UPR). Una, ang chaperone BIP gibuhian gikan sa intraluminal domain sa ER sensors IRE1 ug PERK aron mabugkos ang wala mapilo / sayop nga mga protina. Gitugotan niini ang dimerization ug trans-auto-phosphorylation sa ilang mga cytosolic domain. Ang pagpaaktibo sa PERK moresulta sa direktang NRF2 phosphorylation sa Ser40, nga mosangpot sa NRF2 translocation ngadto sa nucleus ug pagpaaktibo sa target nga mga gene. Ang pagpaaktibo sa IRE1 nag-aghat sa pagrekrut sa TRAF2 nga gisundan sa ASK1 ug JNK phosphorylation ug pagpaaktibo. Ingon nga ang JNK gi-report sa phosphorylate ug pag-activate sa NRF2, makatarunganon nga hunahunaon nga ang pagpaaktibo sa IRE1 magdala ngadto sa dugang nga kalihokan sa NRF2.
Daghang mga pagtuon sa induction sa UPR ang gihimo uban sa inhibitor sa protina glycosylation tunicamycin. Ang NRF2 mopatim-aw nga kinahanglanon alang sa pagpugong sa tunicamycin-induced apoptotic cell death [31] ug ang pagpaaktibo niini ubos niini nga mga kondisyon gimaneho sa autophagic degradation sa KEAP1 [51]. Tungod niini, ang shRNA-mediated silencing sa NRF2 nga ekspresyon sa ?TC-6 nga mga selula, usa ka murine insulinoma ?-cell line, kamahinungdanon nagdugang sa tunicamycin-induced cytotoxicity ug mitultol sa usa ka pagtaas sa ekspresyon sa pro-apoptotic ER stress marker CHOP10. Sa laing bahin, ang pagpaaktibo sa NRF2 pinaagi sa 1,2-dithiole-3-thione (D3T) nagpamenos sa tunicamycin cytotoxicity ug nagpahinay sa ekspresyon sa CHOP10 ug PERK [52]. Makapainteres, ang mga olfactory neuron nga gisumite sa systemic nga aplikasyon sa tunicamycin nagdugang sa NRF2 nga managsama sa ubang mga miyembro sa UPR sama sa CHOP, BIP, XBP1 [53]. Kini nga mga resulta gipalapdan ngadto sa mga pagtuon sa vivo, ingon nga lateral ventricular infusion sa tunicamycin sa mga ilaga nga gipahinabo nga ekspresyon sa PERK ug NRF2 sa hippocampus nga giubanan sa mahinungdanon nga mga kakulangan sa panghunahuna, nadugangan ang TAU phosphorylation ug A?42 nga mga deposito [54].
Ang NRF2 Up-Regulates Key Genes alang sa Maintenance sa ER Physiology
Ang ER lumen nagkinahanglan og daghang suplay sa GSH gikan sa cytosol aron mapadayon ang disulfide chemistry. Ang NRF2 modulates sa mahinungdanong mga enzyme sa GSH metabolismo sa utok, sama sa cystine/glutamate transport, ?-glutamate cysteine synthetase (?-GS), glutamate-cysteine ligase catalytic ug modulator subunits (GCLC ug GCLM), glutathione reductase (GR) ug glutathione peroxidase (GPX) (gisusi sa [55]). Ang kalabutan sa NRF2 sa pagpadayon sa GSH sa ER gisuportahan sa pagpangita nga ang pharmacological o genetic nga pagpaaktibo sa NRF2 nagresulta sa dugang nga GSH synthesis pinaagi sa GCLC / GCLM, samtang ang pagpugong sa pagpahayag niini nga mga enzyme pinaagi sa NRF2-knockdown hinungdan sa usa ka panagtapok sa nadaot. protina sa sulod sa ER padulong sa pagpaaktibo sa UPR [56].
Sa C. elegans daghang mga sangkap sa UPR target nga mga gene nga gi-regulate sa SKN-1, lakip ang Ire1, Xbp1 ug Atf6. Bisan kung ang NRF2 nag-regulate sa pagpahayag sa daghang peroxidase (PRX) ug glutathione peroxidase (GPX) nga mga gene sa mga mammal (gisusi sa [57]), ang GPX8 lamang ang usa ka bona fide nga ER-localized nga enzyme, nga nagtago sa KDEL retrieval signal [58]. Ang pagkawala sa GPX8 hinungdan sa pagpaaktibo sa UPR, pagtulo sa ERO1?-nakuha nga hydrogen peroxide sa cytosol ug pagkamatay sa cell. Hydrogen peroxide nga nakuha gikan sa ERO1? Ang kalihokan dili magkatag gikan sa ER ngadto sa cytosol tungod sa hiniusang aksyon sa GPX8 ug PRX4 [59]. Niining bahina, ang usa ka pagtuki sa antioxidant defense pathway-gene expression array gamit ang RNA gikan sa wild type ug NRF2-null nga tisyu sa mga ilaga, nagpadayag nga ang ekspresyon sa GPX8 gipaubos-regulate sa pagkawala sa NRF2 [60]. Nahiuyon niini, ang pag-analisa sa transcriptome gikan sa mga sample sa pasyente nga nag-antos sa myeloproliferative neoplasms, polycythemia o myelofibrosis, mga sakit nga nakig-uban usab sa oxidative stress ug low-grade nga chronic nga panghubag, nagpakita sa ubos nga lebel sa ekspresyon sa NRF2 ug GPX8 kumpara sa kontrol nga mga subject [61]. Wala pa'y mga pagtuon nga espesipikong naglambigit sa GPX8 sa proteksyon sa utok sa tawo apan ang usa ka transcriptome analysis sa mga ilaga nagpakita sa usa ka compensatory nga pagtaas sa GPX8 isip tubag sa Parkinsonian toxin MPTP [62].
Epekto sa NRF2 sa UPR Dysregulation sa Neurodegenerative Diseases
Ang malfunction sa mga enzyme sa PDI ug ang kanunay nga pagpaaktibo sa UPR mahimo’g magsugod o makapadali sa neurodegeneration. Ang mga neuron nga naapektuhan sa sakit, mga modelo sa hayop sa sakit nga neurodegenerative ingon man ang mga tisyu sa post-mortem sa tawo nagpamatuod sa pag-regulate sa daghang mga UPR-marker sa kadaghanan niini nga mga sakit. Ang pagbag-o sa agianan sa PDI / UPR sa mga sakit nga neurodegenerative maayo nga gisusi sa [63] apan ang mga mosunud nga mga highlight gikan sa mga sample sa post-mortem sa utok kinahanglan nga tagdon. Ang lebel sa PDI nadugangan sa tangle-bearing neurons ug sa Lewy Bodies sa AD ug PD nga mga pasyente, matag usa [64], [65]. Ang PDI ug ERP57 gi-regulate sa CSF gikan sa mga pasyente sa ALS ug sa utok gikan sa mga subject sa CJD [66], [67], [68]. Ang BIP, PERK, IRE1 ug ATF6 gipataas sa mga sample gikan sa mga pasyente nga adunay AD, PD o ALS [69], [70], [71], [67]. Ang BIP, CHOP ug XBP1 gipataas sa post-mortem nga mga sampol sa utok gikan sa HD [72], [73]. Dugang pa, ang up-regulation sa ERP57, GRP94 ug BIP nakit-an sa mga tisyu sa cortex gikan sa mga pasyente sa CJD [74]. Sa kinatibuk-an, kini nga ebidensya nagpadayag nga ang akumulasyon sa sayop nga pagkapilo sa mga protina sa parenchyma sa utok mosangpot sa usa ka makadaot ug laygay nga pagpaaktibo sa UPR. Makapainteres, adunay bag-o nga pagtuon nga nagsumpay sa pagpaaktibo sa NRF2 sa PERK sa sayong bahin sa AD. Sa kini nga pagtuon, gisusi sa mga tigsulat kung ang oxidative stress nagpataliwala sa mga pagbag-o sa NRF2 ug ang UPR mahimo’g naglangkob sa sayo nga mga panghitabo sa AD pathogenesis pinaagi sa paggamit sa mga selula sa dugo sa peripheral sa tawo ug usa ka modelo sa AD transgenic mouse sa lainlaing mga yugto sa sakit. Ang dugang nga oxidative stress ug dugang pSer40-NRF2 naobserbahan sa peripheral blood mononuclear cells sa tawo nga nahimulag gikan sa mga indibidwal nga adunay malumo nga pagkadaot sa panghunahuna. Dugang pa, gi-report nila ang ningdaot nga ER calcium homeostasis ug gi-regulate nga ER-stress marker sa kini nga mga selyula gikan sa mga indibidwal nga adunay malumo nga pagkadaot sa panghunahuna ug malumo nga AD [75].
Mutual Regulation sa NRF2 ug ang Ubiquitin Proteasome System (UPS)
Gibag-o sa UPS ang mga lebel sa protina sa NRF2
Ang UPS miapil sa pagkadaot sa mga nadaot o sayop nga pagkapilo nga mga protina ug nagkontrol sa lebel sa mga importanteng molekula sa regulasyon sa cytosol ug sa nucleus. Ang sentro nga kinauyokan niini nga sistema kay usa ka dako nga multisubunit nga enzyme nga adunay sulod nga proteolytic active complex nga ginganlag 20S. Ang 20S nga kinauyokan nga proteasome nagdaot sa gibuklad nga mga protina, apan ang pagbugkos sa lainlaing mga regulatory protein complex nagbag-o sa espesipiko ug kalihokan sa substrate. Pananglitan, ang pagdugang sa usa o duha ka 19S regulatory subunits sa 20S core naglangkob sa 26S proteasome ug nagbag-o sa espesipiko niini ngadto sa lumad nga napilo nga mga protina [76], [77]. Ang pagkadaot sa proteasomal nagkinahanglan og covalent binding sa ubiquitin. Ang conjugation sa ubiquitin nagpadayon pinaagi sa tulo ka lakang nga mekanismo sa cascade. Una, ang ubiquitin-activating enzyme E1 nagpalihok sa ubiquitin sa usa ka ATP-required nga reaksyon. Dayon, ang usa ka E2 enzyme (ubiquitin-carrier protein o ubiquitin-conjugating enzyme) mobalhin sa activated ubiquitin gikan sa E1 ngadto sa substrate nga espesipikong gigapos sa usa ka sakop sa ubiquitin-protein ligase nga pamilya, nga ginganlag E3. Bisan kung ang eksaktong kapalaran sa ubiquitinated-protein magdepende sa kinaiya sa ubiquitin chain, kini nga proseso sa kasagaran moresulta sa pagkadaot sa 26S proteasome [78].
Ang E3-ligase KEAP1 mao ang labing nailhan nga tigpugong sa NRF2. Ang mekanismo sa regulasyon sa KEAP1 elegante nga nagpatin-aw kung giunsa ang pag-adjust sa lebel sa NRF2 sa pag-usab-usab sa oxidant. Ubos sa basal nga mga kondisyon, ang bag-ong gi-synthesize nga NRF2 gikuha sa homodimer KEAP1, nga nagbugkos sa usa ka NRF2 nga molekula sa duha ka amino acid sequences nga adunay ubos (aspartate, leucine, glycine; DLG) ug taas (glutamate, threonine, glycine, glutamate; ETGE) affinity. Ang interaksyon sa KEAP1 nagtabang sa pagpresentar sa NRF2 sa CULLIN3 / RBX1 protein complex, nga miresulta sa ubiquitination niini ug sa sunod nga proteasomal degradation. Bisan pa, ang pagbag-o sa redox sa KEAP1 nagpugong sa pagpresentar sa NRF2 sa UPS nga girepresentahan sa CULLIN3 / RBX1. Ingon usa ka sangputanan, ang bag-ong gi-synthetized nga NRF2 nakaikyas sa pagkadaot nga nagsalig sa KEAP1, natipon sa nucleus ug gi-aktibo ang mga gene nga adunay ARE [79], [80], [81], [82].
Ang E3-ligase adapter ?-TrCP usa usab ka homodimer nga miapil sa mga panghitabo sa pagsenyas nga may kalabutan sa phosphorylation sa NRF2 sa GSK-3?. Kini nga kinase phosphorylates piho nga serine residues sa NRF2 (aspartate, serine, glycine, isoleucine serine; DSGIS) sa paghimo sa usa ka degradation domain nga unya giila sa ?-TrCP ug tagged proteasome degradation sa usa ka CULLIN1/RBX1 complex. Ang pag-ila sa piho nga amino acids nga phosphorylated sa GSK-3? sa niini nga degron gipahigayon pinaagi sa usa ka kombinasyon sa site-directed mutagenesis sa Neh6 domain, 2D-gel electrophoresis [15], [26] ug mass spectroscopy [83]. Tungod niini, ang pagdili sa GSK-3? pinaagi sa labi ka pinili nga mga tambal o siRNA batok sa GSK-3 isoforms miresulta sa pagtaas sa lebel sa protina sa NRF2. Ang susamang mga resulta nakit-an sa mga siRNA batok sa ?-TrCP isoforms 1 ug 2. Pagpalig-on sa NRF2 nga nagsunod sa GSK-3? Ang pagpugong nahitabo sa KEAP1-kulang nga mouse embryo fibroblasts ug sa usa ka ectopically nga gipahayag nga NRF2 nga pagtangtang mutant nga kulang sa kritikal nga mga residu sa ETGE alang sa high-affinity binding sa KEAP1, dugang nga nagpakita sa usa ka KEAP1-independent nga regulasyon.
Sa konteksto sa mga sakit nga neurodegenerative, atong mahanduraw ang modulasyon sa NRF2 sa UPS sa duha ka lainlaing paagi. Sa usa ka bahin, ang sistema sa KEAP1 makamatikod sa redox imbalance nga nakuha gikan sa misfolded protein accumulation, samtang ang GSK-3/?-TrCP axis maglihok isip aktibong partisipante sa signaling transduction nga giusab sa pagkawala sa proteostasis (Fig. 2).
Figure 2 Ang UPS hugot nga nagkontrol sa lebel sa NRF2. Ubos sa mga kondisyon sa homeostatic, ang ubos nga lebel sa NRF2 gipadayon pinaagi sa aksyon sa E3 ligases adapters KEAP1 ug ?-TrCP. Sa wala, ang NRF2 nagbugkos sa mga domain sa Kelch sa usa ka KEAP1 homodimer pinaagi sa usa ka ubos nga (DLG) ug usa ka taas nga (ETGE) nga mga motibo sa affinity. Pinaagi sa BTB domain niini, ang KEAP1 dungan nga nagbugkos sa usa ka CULLIN3 / RBX1 complex, nga nakapahimo sa NRF2 ubiquitination ug degradation sa 26 S proteasome. Dugang pa, GSK-3? phosphorylates Ser335 ug Ser338 residues sa NRF2 sa paghimo sa usa ka degradation domain (DpSGIpSL) nga unya giila sa ubiquitin ligase adapter ?-TrCP ug gi-tag alang sa proteasome degradation sa usa ka CULLIN3/RBX1 complex. Husto, Sa pagkaladlad sa mga reaktibo nga espisye sa oksiheno o mga electrophile nga kritikal nga Cys residues sa KEAP1 giusab, nga naghimo sa KEAP1 nga dili maka-interact nga episyente sa NRF2 o CULLIN3 / RBX1 ug unya kini nga transcription factor nagdugang sa iyang half-life ug transcriptional nga kalihokan ngadto sa ARE-genes. Ang mga agianan sa pagsenyas nga moresulta sa pagdili sa GSK-3?, ang ingon nga AKT phosphorylation sa Ser9, moresulta sa pagkadaot sa NRF2 pinaagi sa proteasome, akumulasyon ug induction sa mga target nga gene.
Ang NRF2 Nagpataas sa Kalihokan sa UPS Pinaagi sa Transcriptional Control sa Proteasome Subunits
Ang NRF2 nag-regulate sa pagpahayag sa daghang mga subunit sa proteasome, sa ingon nanalipod sa selula gikan sa pagtipon sa makahilo nga mga protina. Kaluhaan ka proteasome- ug ubiquitination-related nga mga gene makita nga gi-regulate sa NRF2, sumala sa usa ka halapad nga microarray analysis gikan sa atay RNA nga gi-set up sa NRF2 inducer D3T [84]. Sa usa ka posterior nga pagtuon, ang parehas nga mga awtor nagpamatuod nga ang ekspresyon sa kadaghanan nga mga subunit sa 26S proteasome gipauswag hangtod sa tulo ka pilo sa mga atay gikan sa mga ilaga nga gitambalan sa D3T. Ang lebel sa protina sa subunit ug kalihokan sa proteasome koordinado nga nadugangan. Bisan pa, walay induction nga nakita sa mga ilaga diin ang transcription factor NRF2 nabalda. Ang kalihokan sa tigpasiugda sa PSMB5 (20S) proteasome subunit miuswag uban sa NRF2 overexpression o pagtambal sa mga activator sa mouse embryonic fibroblasts, ug ang mga ARE giila sa proximal promoter sa PSMB5 [85]. Ang pagpaaktibo sa pharmacological sa NRF2 miresulta sa taas nga lebel sa ekspresyon sa representante nga proteasome subunits (PSMA3, PSMA6, PSMB1 ug PSMB5) lamang sa non-senescent human fibroblasts nga adunay functional NRF2 [86]. Ang pagpaaktibo sa NRF2 sa panahon sa pagpahiangay sa oxidative stress nagresulta sa taas nga pagpahayag sa PSMB1 (20S) ug PA28? subunits (o S11, proteasome regulator) [87]. Dugang pa, ang mga resulta gikan sa human embryonic stem cell nagpadayag nga ang NRF2 nagkontrol sa ekspresyon sa proteasome maturation protein (POMP), usa ka proteasome chaperone, nga sa baylo modulates sa pagdaghan sa mga self-renewing human embryonic stem cells, tulo ka germ layer differentiation ug cellular reprogramming. 88]. Tanan, kini nga mga pagtuon nagpakita nga ang NRF2 nag-regulate sa pagpahayag sa mga hinungdan nga sangkap sa UPS ug busa aktibo nga nakatampo sa clearance sa mga protina nga mahimo’g makahilo.
Ang NRF2-UPS Axis sa Neurodegenerative nga mga Sakit
Ang papel sa UPS sa mga sakit nga neurodegenerative usa ka natad sa intensive debate. Gi-report sa mga pasiunang pagtuon ang pagkunhod sa kalihokan sa proteasome sa mga necropsies sa tawo sa mga pasyente nga naapektuhan sa daghang mga sakit nga neurodegenerative. Bisan pa, ang ubang mga pagtuon nga naggamit sa in vitro ug in vivo nga mga pamaagi nakit-an nga wala nagbag-o o bisan ang pagtaas sa kalihokan sa proteasome (gisusi sa [89]). Usa ka posible nga katin-awan alang niini nga kalainan mao nga ang lebel sa mga sangkap sa UPS mahimong mausab sa panahon sa pag-uswag sa sakit ug sa lainlaing mga rehiyon sa utok sama sa gisugyot alang sa mga target sa NRF2.
Bisan pa niini nga kontrobersiya, kinahanglan nga matikdan nga ang up-regulation sa ARE-containing proteasome genes magpalig-on sa UPS pinaagi sa pagdugang sa clearance sa makahilong mga protina sa utok. Sa tinuud, ang ablation sa NRF1, usab modulator sa pagtubag sa antioxidant, sa mga selula sa neuronal hinungdan sa ningdaot nga kalihokan sa proteasome ug neurodegeneration. Ang mga eksperimento sa immunoprecipitation sa Chromatin ug pagtuki sa transkripsyon nagpakita nga ang PSMB6 gi-regulate sa NRF1. Dugang pa, ang gene expression profiling misangpot sa pag-ila sa NRF1 isip usa ka importante nga transcriptional regulator sa proteasome genes sa mga neuron, nga nagsugyot nga ang mga perturbation sa NRF1 mahimong makatampo sa pathogenesis sa neurodegenerative nga mga sakit [90]. Makapainteres, ang NRF1 ug ang taas nga isoform niini nga gitawag nga TCF11 gipakita nga nag-regulate sa ARE nga adunay sulud nga proteasome nga mga gene sa pagdili sa proteasome sa usa ka feedback loop aron mabayran ang pagkunhod sa kalihokan sa proteolytic [91], [92].
Mahitungod sa NRF2, adunay usa ka correlation sa pagkunhod sa NRF2, RPT6 (19 S) ug PSMB5 (20 S) nga lebel sa midbrain sa DJ-1-kulang nga mga ilaga nga gitambalan sa neurotoxin paraquat [93]. Dugang pa, ang natural nga nahitabo nga compound sulforaphane (SFN) naghatag usa ka labi ka lig-on nga imahe sa NRF2 ingon usa ka hinungdanon nga modulator sa UPS. Ang mga eksperimento sa vitro nga adunay murine neuroblastoma Neuro2A nga mga selyula nagpamatuod sa usa ka gipauswag nga pagpahayag sa mga catalytic subunits sa proteasome, ingon man ang mga kalihokan sa peptidase niini agig tubag sa SFN. Kini nga tambal nanalipod sa mga selula gikan sa hydrogen peroxide-mediated cytotoxicity ug protina oxidation sa paagi nga nagsalig sa proteasome function [94]. Dugang pa, si Liu ug mga kauban sa trabaho migamit ug reporter mouse aron mamonitor ang kalihokan sa UPS agig tubag sa SFN sa utok. Kini nga mga ilaga kanunay nga nagpahayag sa berde nga fluorescence protein (GFP) nga gisagol sa usa ka constitutive degradation signal nga nagpasiugda sa paspas nga pagkadaot sa UPS (GFPu). Sa cerebral cortex, gipaubos sa SFN ang lebel sa GFPu nga adunay managsama nga pagtaas sa chymotrypsin-like (PSMB5), caspase-like (PSMB2), ug trypsin-like (PSMB1) nga mga kalihokan sa 20 S proteasome. Dugang pa, ang pagtambal sa mga selula nga nakuha sa Huntington nga adunay SFN nagpadayag nga ang pagpaaktibo sa NRF2 nagpauswag sa pagkadaot sa mHtt ug pagkunhod sa cytotoxicity sa mHtt [95]. Ang mayor nga mekanismo sa aksyon sa SFN mao ang pinaagi sa induction sa NRF2 [96]. Ang espesipikong kontribusyon sa NRF2 kinahanglan nga hisgutan gamit ang NRF2-null nga mga sistema sa dugang nga mga pagtuon.
Functional nga Koneksyon Tali sa NRF2 ug Macroautophagy
Ang NRF2 Protein Levels kay Modulated sa Adapter Protein P62
Ang autophagy nagtumong sa pagkadaot sa cytosolic nga mga sangkap sulod sa lysosomes. Kini nga proseso gigamit alang sa paghawan sa dugay na nga kinabuhi ug sayop nga pagkapilo nga mga protina ingon man sa nadaot nga mga organel. Ang usa ka direkta nga sumpay tali sa NRF2 ug autophagy unang naobserbahan sa koneksyon sa adapter protein p62, gitawag usab nga SQSTM1 [97], [98], [99], [100], [101]. Kini nga protina nag-shuttle sa ubiquitinated nga mga protina ngadto sa proteasomal ug lysosomal nga mga makinarya sa degradasyon ug mga sequester nga nadaot ang mga protina ngadto sa mga aggregate sa wala pa ang ilang pagkadaut. Ang P62 nagpresentar sa usa ka ubiquitin-associated (UBA) domain, alang sa pagbugkos sa ubiquitinated proteins, ug usa ka LC3-interacting region (LIR) alang sa pag-integrate sa autophagosomal membrane pinaagi sa autophagy receptor LC3.
Bisan tuod ang p62-mediated induction sa NRF2 ug ang target nga mga gene niini unang gitaho niadtong 2007 [102], ang mekanismo sa molekula wala pa hingpit nga nasabtan hangtud sa pagkadiskobre sa interaksyon niini sa KEAP1 [103], [98], [99], [100]. ], [101]. Giila ni Komatsu ug mga kauban sa trabaho ang usa ka KEAP1 interacting region (KIR) sa p62 nga naggapos sa KEAP1 sa samang basic surface pocket sama sa NRF2 ug adunay usa ka binding affinity susama sa ETGE motif sa NRF2, nga nagsugyot sa kompetisyon tali sa p62 ug NRF2. Ang phosphorylation sa Ser351 sa KIR motif sa p62 (349-DPSTGE-354) gipakita aron madugangan ang pagkadugtong niini alang sa KEAP1, nga nakigkompetensya sa NRF2 nga nagbugkos ug nagtugot sa pagtipon niini ug pagpaaktibo sa transkripsyon sa mga target nga gene niini [98], [99]. Sa tinuud, ang sobrang pagpahayag sa p62 misangpot sa pagkunhod sa NRF2 ubiquitination ug ang sangputanan nga pag-stabilize ingon man ang pag-induction sa mga target nga gene niini [104]. Ang ubang mga kinase gisugyot nga moapil sa p62 phosphorylation. Ang target sa mammalian sa rapamycin complex 1 (mTORC1) mahimong iapil, tungod kay ang pagtambal sa mTOR inhibitor nga rapamycin nagpugong sa phosphorylation sa p62 ug ang down-regulation sa KEAP1 sa pagtambal sa arsenite. Bag-ohay lang, gipakita nga ang TGF-?-activate kinase 1 (TAK1) mahimo usab nga phosphorylate p62, pagpausbaw sa KEAP1 degradation ug NRF2-up-regulation. Ang mga tigsulat niini nga pagtuon nagsugyot nga kini usa ka paagi sa pag-regulate sa cellular redoxtasis ubos sa steady-state nga mga kondisyon, tungod kay ang TAK1-kakulangan nag-regulate sa ROS kung wala ang bisan unsang exogenous oxidant sa lainlaing mga tisyu sa mouse nga susama sa pagkunhod sa lebel sa protina sa NRF2 [105]. ].
Ang usa ka p62 nga pagtukod nga kulang sa UBA domain makahimo gihapon sa pagbugkos sa KEAP1, nga nagpasabot nga ang interaksyon wala magdepende sa ubiquitinated KEAP1 [101]. Bisan pa, ang p62 homologue sa Drosophila melanogaster, nga ginganlag Ref(2), wala’y sulud nga KIR motif ug dili direkta nga nakig-uban sa DmKEAP1, bisan kung mahimo kini magbugkos sa ubiquitinated DmKEAP1 pinaagi sa domain sa UBA. Dugang pa, ang DmKEAP1 mahimong direktang makig-uban sa Atg8 (homologue sa mammalian LC3). Ang kakulangan sa KEAP1 nagresulta sa Atg8 ug autophagy induction nga nagsalig sa NRF2 orthologue CncC ug independente sa TFEB / MITF [106]. Ang relasyon tali sa NRF2 ug autophagy daw gitipigan bisan pa, nga nagpasiugda sa kalambigitan niini.
Ang induction sa NRF2 sa p62 mao ang resulta sa kompetisyon sa paggapos sa KEAP1 ug pagkadaut sa KEAP1 sa lysosome. Ang pagpahilom sa p62 nga adunay siRNA nagdoble sa KEAP1 nga katunga sa kinabuhi nga managsama sa pagkunhod sa NRF2 ug ang target nga mga gene niini [101]. Sa pag-uyon, ang ablation sa p62 nga ekspresyon nagpamatuod sa pagtaas sa lebel sa KEAP1 kumpara sa ihalas nga tipo nga mga ilaga. May kalabotan kaayo, ang pag-uswag sa lebel sa KEAP1 wala maapektuhan sa mga inhibitor sa proteasome apan gipakunhod ubos sa autophagy nga nagpahinabo sa kagutom [107]. Sa tinuud, ang KEAP1 naa sa mga selula sa mammalian sa mga autophagic vesicle nga gidekorasyonan sa p62 ug LC3 [99], [100], [103]. Ang tanan nga kini nga mga datos nagsugyot nga ang KEAP1 usa ka substrate sa makinarya sa macroautophagy, apan kini nga isyu kinahanglan nga analisahon nga mas detalyado tungod sa paglungtad sa pipila ka mga kontrobersyal nga mga sangputanan. Ang lebel sa protina sa KEAP1 nadugangan sa Atg7-null nga mga ilaga, usa ka mahinungdanong effector sa macroautophagy [107], apan ang pharmacological inhibition sa macroautophagy nga adunay torin1, E64 / pepstatin o bafilomycin napakyas sa pagtigum sa KEAP1 [107], [100]. Sa kinatibuk-an, kini nga mga resulta nagsugyot nga ang pagtaas sa lebel sa p62 nag-sequester sa KEAP1 ngadto sa autophagic vacuoles ug tingali kini nga mga resulta sa KEAP1 autophagic degradation nga nagtugot sa NRF2 activation (Fig. 3). Duha ka lainlaing mga pagtuon ang nagtaho nga ang sulfinic acid reductases SESTRINS adunay hinungdanon nga papel sa kini nga konteksto. Ang SESTRIN 2 nakig-uban sa p62, KEAP1 ug RBX1 ug nagpadali sa p62-dependent nga pagkadaot sa KEAP1 ug NRF2 nga pagpaaktibo sa mga target nga gene [108]. Ang laing pagtuon nagpakita nga ang SESTRIN 2 nakig-interact sa ULK1 ug p62, nga nagpasiugda sa phosphorylation sa p62 sa Ser403 nga nagpadali sa pagkadaut sa mga protina sa kargamento lakip na ang KEAP1 [109].
Ang Figure 3 nga lebel sa NRF2 gi-regulate sa adapter protein p62. Ang phosphorylation sa Ser 351 sa KIR motif sa p62 (349-DPSTGE-354) sa mTORC1, TAK1 o uban pang kinases moresulta sa dugang nga affinity alang sa pagbugkos sa KEAP1 tungod sa pagkasama sa ETGE motif sa NRF2. Ingon usa ka sangputanan, ang phosphorylated p62 nagbalhin sa NRF2 ug nagbugkos sa KEAP1. Ang LIR motif sa p62 makahimo sa interaksyon sa LC3 sa autophagosomal membrane, aron ang p62-KEAP1 complex sa kadugayan madaot sa lysosome. Ingon usa ka sangputanan ang NRF2 makahimo sa pagtipon, pagbalhin sa nucleus ug pagdugang sa transkripsyon sa mga gene nga adunay ARE, lakip ang p62. Kini nga mekanismo sa regulasyon naghatag usa ka kanunay nga tubag sa NRF2, tungod kay ang KEAP1 kinahanglan nga bag-o nga gi-synthesize aron mapugngan ang kalihokan sa NRF2.
Modulasyon sa Macroautophagy Genes pinaagi sa NRF2
Ang NRF2 nag-regulate sa pagpahayag sa mga may kalabutan nga mga gene alang sa macroautophagy ingon man kini alang sa UPR ug sa UPS. Ang una nga ebidensya naggikan sa mga pagtuon diin ang p62 nga ekspresyon gipakita nga naaghat sa pagkaladlad sa mga electrophile, ROS ug nitric oxide [110], [111], [112]. Ang mekanismo sa induction gihulagway pipila ka tuig ang milabay uban sa pagpangita nga ang p62 naglangkob sa usa ka functional ARE sa iyang gene promoter [99]. Sa usa ka bag-o nga pagtuon, daghang uban pang mga functional ARE ang nakit-an ug gi-validate pagkahuman sa bioinformatics analysis ug ChIP assays. Dugang pa, ang mouse embryonic fibroblasts ug cortical neurons gikan sa Nrf2-knockout nga mga ilaga nagpakita sa pagkunhod sa p62 nga ekspresyon, nga mahimong maluwas sa usa ka NRF2-expressing lentivirus. Sa susama, ang kakulangan sa NRF2 mikunhod sa lebel sa p62 sa nasamdan nga mga neuron gikan sa mga ilaga nga hippocampus [36]. Busa, gisugyot nga ang pagpaaktibo sa NRF2 nagdugang sa lebel sa p62, nga miresulta sa pagkadaut sa KEAP1 ug gipaboran ang dugang nga pag-stabilize sa NRF2 sa usa ka positibo nga feedback loop. Kini nga non-canonical nga mekanismo sa NRF2 induction nanginahanglan mga pagbag-o sa ekspresyon sa gene ug mahimong usa ka may kalabutan nga tubag sa dugay nga cellular stress.
Ang cargo recognition protein NDP52 gipakita nga transcriptionally regulated sa NRF2. Ang NDP52 nagtrabaho sa susama nga paagi sa p62, nga nag-ila sa ubiquitinated nga mga protina ug nakig-uban sa LC3 pinaagi sa LIR domain, aron ang mga kargamento madaot sa lysosomes. Lima ka putative ARE ang nakit-an sa Ndp52 promoter DNA sequence. Tulo niini giila nga adunay lainlaing mutant constructs ug ChIP assays nga kinahanglanon alang sa NRF2-mediated Ndp52 transcription [113]. Sa nota, ang mga lebel sa Ndp52 mRNA mikunhod sa hippocampus sa Nrf2-knockout nga mga ilaga. Ang usa niini nga mga han-ay gipamatud-an usab sa usa ka independenteng pagtuon isip usa ka NRF2-regulated ARE [36].
Bisan pa, ang papel sa NRF2 sa modulasyon sa autophagy dili limitado sa induksiyon niining duha ka mga protina sa pag-ila sa kargamento. Aron makakuha og mas lawom nga pagsabot sa papel sa NRF2 sa modulasyon sa dugang nga mga gene nga may kalabutan sa autophagy, gisusi sa among grupo ang chromatin immunoprecipitation database ENCODE alang sa duha ka protina, MAFK ug BACH1, nga nagbugkos sa NRF2-regulated AREs. Gamit ang usa ka script nga namugna gikan sa JASPAR's consensus ARE sequence, nahibal-an namon ang daghang mga putative ARE sa daghang mga autophagy genes. Napulog-duha niini nga mga han-ay ang gi-validate isip NRF2 regulated AREs sa siyam ka autophagy genes, kansang ekspresyon mikunhod sa mouse embryo fibroblasts sa Nrf2-knockout nga mga ilaga apan mahimong ibalik sa usa ka NRF2-expressing lentivirus. Gipakita sa among pagtuon nga ang NRF2 nag-aktibo sa pagpahayag sa pipila nga mga gene nga nalambigit sa lainlaing mga lakang sa proseso sa autophagic, lakip ang pagsugod sa autophagy (ULK1), pag-ila sa kargamento (p62 ug NDP52), pagporma sa autophagosome (ATG4D, ATG7 ug GABARAPL1), pagpalawig (ATG2B ug ATG5). ), ug autolysosome clearance (ATG4D). Tungod niini, ang autophagy flux agig tubag sa hydrogen peroxide nadaot sa dihang wala ang NRF2 [36].
Kalambigitan sa NRF2-Mediated Macroautophagy Genes Expression sa Neurodegenerative Disorders
Ang depekto nga autophagy gipakita nga adunay hinungdanon nga papel sa daghang mga sakit nga neurodegenerative [114] ug ang ablation sa autophagy nagdala sa neurodegeneration sa mga ilaga [115], [116]. Gipadayag sa Atg7-knockout nga mga ilaga nga ang kakulangan sa autophagy nagresulta sa pagtipon sa p62 sa mga ubiquitin-positibo nga paglakip nga mga lawas. Ang KEAP1 gi-sequester niining mga inclusion nga mga lawas, nga mitultol sa NRF2 stabilization ug induction sa target genes [103]. Importante, ang sobra nga akumulasyon sa p62 uban sa ubiquitinated nga mga protina giila sa mga sakit nga neurodegenerative, lakip ang AD, PD ug ALS [117]. Sa pagkatinuod, ang mga neuron nga nagpahayag sa taas nga lebel sa APP o TAU sa mga pasyente sa AD nagpahayag usab sa p62 ug nukleyar nga NRF2, nga nagsugyot sa ilang pagsulay sa pagpaubos sa intraneuronal aggregates pinaagi sa autophagy [36].
Ang kakulangan sa NRF2 nagpasamot sa panagsama sa protina sa konteksto sa AD. Sa tinuud, ang pagtaas sa lebel sa phosphorylated ug sarkosyl-insoluble nga TAU makit-an sa Nrf2-knockout nga mga ilaga, bisan kung wala’y kalainan sa mga kalihokan sa kinase o phosphatase nga makit-an kung itandi sa wild-type nga background [113]. Importante, ang NDP52 gipakita nga co-localize sa TAU sa murine neurons ug direktang interaksyon tali sa phospho-TAU ug NDP52 gipakita sa mga eksperimento sa co-immunoprecipitation sa mga ilaga ug AD sample, nga nagpunting sa papel niini sa TAU degradation. Makaiikag, ang pagpahilom sa NDP52, p62 o NRF2 sa mga neuron miresulta sa dugang nga phospho-TAU [113], [118]. Dugang pa, ang dugang nga intraneuronal nga mga aggregate sa APP nakit-an sa hippocampus sa APP / PS1? E9 nga mga ilaga kung wala ang NRF2. Nalambigit kini sa giusab nga mga marker sa autophagy, lakip ang nadugangan nga phospho-mTOR/mTOR ug phospho-p70S6k/p70S6k ratios (nagpaila sa autophagy inhibition), gipadako nga lebel sa pre-cathepsin D ug mas daghang gidaghanon sa mga multivesicular nga lawas [119]. Sa mga ilaga nga nagpahayag sa tawhanong APP (V717I) ug TAU (P301L), ang kakulangan sa NRF2 misangpot sa pagtaas sa lebel sa total ug phospho-TAU sa dili matunaw nga tipik ug nagdugang sa intraneuronal APP aggregates, uban sa pagkunhod sa neuronal nga lebel sa p62, NDP52, ULK1, ATG5 ug GABARAPL1. Ang co-localization tali sa adapter protein p62 ug APP o TAU gikunhoran kung wala ang NRF2 [36]. Sa kinatibuk-an, kini nga mga resulta nagpasiugda sa kamahinungdanon sa NRF2 sa neuronal autophagy.
Lainlaing Transcription Factors Act Coordinately to Modulate Proteostasis
Ubos sa makanunayon nga kahimtang sa kahimtang, ang proteostasis gikontrol pinaagi sa mga interaksyon sa protina-protina ug mga pagbag-o pagkahuman sa paghubad nga nakakuha usa ka paspas nga tubag. Bisan pa, ang pagpahiangay sa cellular nanginahanglan sa regulasyon sa transkripsyon sa UPR, UPS ug mga autophagy nga mga gene. Sa pagkonsiderar nga ang mga selula sa nerbiyos padayon nga gisumite sa ubos nga grado nga makahilo nga mga insulto, lakip ang oxidative ug proteotoxic nga stress, ang usa ka pagpalig-on sa proteostasis nga gipahinabo sa transcriptional modulation mahimong makatabang sa pagpugong sa pagkadaot sa utok.
Sa kaso sa UPR, ang pagpaaktibo sa matag usa sa tulo ka mga bukton sa katapusan moresulta sa transcriptional induction sa pipila ka mga gene (gisusi sa [43]). Pananglitan, ang ATF6-derived fragment (ATF6f) nagbugkos sa ER-stress response elements (ERSE) ug nag-aghat sa pagpahayag sa daghang mga gene, lakip ang XBPI, BIP ug CHOP. Dugang pa, ang PERK signaling mosangpot sa pagpaaktibo sa transcription factor ATF4, nga nagkontrol sa pagpahayag sa daghang UPR-related nga mga gene ug uban pa lakip ang NRF2 target genes Hmox1 ug p62. Sa katapusan, ang pagpaaktibo sa IRE1 nagresulta sa henerasyon sa usa ka aktibo nga transcription factor, gidugtong nga XBP1 (XBP1s), nga nagkontrol sa transkripsyon sa mga gene nga nag-encode sa mga protina nga nahilambigit sa pagpilo sa protina.
Sa laing bahin, ang NRF1 gipakita nga gikinahanglan alang sa proteasomal gene nga ekspresyon sa utok, tungod kay ang Nrf1-knockout nga mga ilaga nagpakita sa pagkunhod sa ekspresyon sa mga gene nga nag-encode sa nagkalain-laing mga subunit sa 20S core, ingon man ang 19S regulatory complex uban sa ningdaot nga proteasomal function [90]. ]. Ang NRF1 ug NRF2 nagbugkos sa mga han-ay sa ARE sa mga rehiyon sa promoter sa mga target nga gene niini, nga nagsugyot nga sila adunay nagsapaw-sapaw nga mga kalihokan sa transkripsyon, bisan kung kini magkalahi sa ilang mga mekanismo sa regulasyon ug cellular localization [120].
Ang mga hinungdan sa transkripsyon sa Forkhead box O (FOXO) nga pamilya nagkontrol sa pagpahayag sa daghang mga gene nga may kalabotan sa autophagy. Sama sa nahitabo sa NRF2, adunay daghang mga lut-od sa regulasyon sa kalihokan sa mga miyembro sa FOXO, nga mahimong maaghat sa nutritional o oxidative stress [121]. Sa katapusan, ang transcription factor nga TFEB, nga gikonsiderar nga master regulator sa lysosomal biogenesis, adunay hinungdanon nga papel sa regulasyon sa autophagy ubos sa kahimtang sa stress sa nutrisyon. Busa, ang pagdili sa mTORC1 mosangpot sa nukleyar nga translokasyon sa TFEB ug induction sa ekspresyon sa autophagy genes [122].
Sa kinatibuk-an, ang paglungtad sa lain-laing mga transcriptional regulators niini nga mga makinarya usab nagsugyot sa crosstalk ug partially redundant nga mga mekanismo nga mahimong makasiguro sa proteostasis ubos sa lain-laing mga kahimtang. Tungod niini, ang NRF2 mahimong adunay kalabutan nga papel sa mga tisyu nga nagsuporta sa taas nga lebel sa stress sa oxidative. Pananglitan, ang oxidative stress-induced NRF2 mahimong molihok ubos sa nutrient-rich nga mga kondisyon aron transcriptionally up-regulate ang autophagy, susama sa nakit-an alang sa TFEB ubos sa mga kondisyon sa kagutom. Dugang pa, ang utok naglihok sa kadaghanan ubos sa mga kondisyon nga puno sa sustansya, nga nagpakita sa NRF2 isip usa ka may kalabutan nga mekanismo aron ma-activate ang autophagy sa mga neuron.
Nagsaad nga Therapeutic Potential alang sa NRF2 sa Proteinopathies
Sa milabay nga pipila ka mga tuig, usa ka dako nga pag-uswag ang nahimo sa kahibalo sa regulatory roles sa UPR, UPS ug autophagy sa NRF2 nga kalihokan, ingon man usab sa reciprocal NRF2-mediated transcription sa mga sangkap niining tulo ka mga sistema. Busa, ang bag-ong mga posibilidad sa terapyutik mahimong motumaw base sa pagpahimulos sa NRF2 ingon usa ka hinungdanon nga regulator sa clearance sa protina sa mga sakit nga neurodegenerative.
Bisan pa, ang usa ka hinungdanon nga nahabilin nga pangutana kung kini mapuslanon o makadaot aron madugangan ang lebel sa NRF2 sa utok. Ang pag-analisa sa epidemiological data mahimong maghatag ug partial nga tubag, tungod kay kini nagpakita nga ang NFE2L2 gene kay polymorphic kaayo ug ang pipila ka single nucleotide polymorphisms nga makita sa promoter niini nga regulatory region mahimong maghatag ug lain-laing �physiological’ variability sa gene expression sa lebel sa populasyon ug pipila ka haplotypes. nalangkit sa pagkunhod sa risgo ug / o nalangan nga pagsugod sa AD, PD o ALS [123]. Dugang pa, sumala sa gihisgutan ni Hayes ug mga kauban [124], ang epekto sa NRF2 mahimong adunay usa ka U-shaped nga tubag, nga nagpasabot nga ang ubos kaayo nga lebel sa NRF2 mahimong moresulta sa pagkawala sa cytoprotection ug dugang nga pagkadaling makuha sa mga stressor, samtang ang sobra nga NRF2 mahimong makabalda sa balanse sa homeostatic ngadto sa usa ka reductive scenario (reductive stress), nga mopabor sa protein misfolding ug aggregation. Ang ubos nga lebel sa NRF2 sa utok nagsuporta sa ideya nga ang usa ka gamay nga regulasyon mahimong igo aron makab-ot ang usa ka kaayohan ubos sa mga kondisyon sa pathological. Sa tinuud, ang proteksiyon nga papel sa pharmacological NRF2-mediated activation sa clearance sa protina gipakita sa lainlaing neurodegeneration cell culture ug sa vivo nga mga modelo.
Ang SFN usa ka pharmacological NRF2 activator nga gipakita aron maaghat ang proteasomal ug autophagy gene expression [95], [36]. Makapainteres, gipakita ni Jo ug mga kauban nga ang SFN nagpamenos sa lebel sa phosphorylated TAU ug nagdugang sa Beclin-1 ug LC3-II, nga nagsugyot nga ang pagpaaktibo sa NRF2 mahimong mapadali ang pagkadaot sa kini nga makahilo nga protina pinaagi sa autophagy [113]. Dugang pa, ang pagkadaut sa mHtt gipalambo sa SFN, ug kini gibalik sa paggamit sa MG132, nga nagpakita sa proteasomal nga pagkadaut niining makahilo nga protina [95]. Ang autophagy-mediated degradation sa phospho- ug insoluble-TAU gitaho sa organic flavonoid fisetin. Kini nga compound nakahimo sa pag-aghat sa autophagy pinaagi sa dungan nga pagpasiugda sa pagpaaktibo ug nukleyar nga paghubad sa TFEB ug NRF2, uban sa pipila sa mga target nga gene niini. Kini nga tubag gipugngan sa TFEB o NRF2 nga pagpahilom [125]. Si Bott ug mga kaubanan nagtaho sa mapuslanong mga epekto sa usa ka dungan nga NRF2, NRF1 ug HSF1 activator sa pagkahilo sa protina sa spinal ug bulbar muscular atrophy, usa ka neurodegenerative disorder tungod sa pagpalapad sa polyglutamine-encoding nga CAG nga gisubli diin ang mga aggregate sa protina anaa [126]. Ang potensyal sa pagpaaktibo sa NRF2 alang sa pagtambal sa mga neurodegenerative disorder gipakita uban ang pag-uyon sa BG-12, ang oral formulation sa NRF2 inducer dimethyl fumarate (DMF), alang sa pagtambal sa multiple sclerosis [127], [128]. Ang kalampusan sa DMF nga adunay mga sakit nga autoimmune nga adunay kusog nga sangkap nga makapahubag nagsugyot nga ang mga sakit nga neurodegenerative mahimong makabenepisyo gikan sa pag-reposition niini nga tambal. Sa usa ka bag-o nga preclinical nga pagtuon sa usa ka ?-synucleinopathy nga modelo sa PD, ang DMF gipakita nga neuroprotective tungod, sa bahin, sa iyang induction sa autophagy [129]. Ang mga pagtuon nga nagreport sa mapuslanon nga mga epekto sa NRF2 sa neurodegeneration apan wala magpunting sa epekto niini sa clearance sa protina labi pa ka daghan (alang sa usa ka komprehensibo nga pagrepaso, tan-awa [7]). Kini adunay kalabutan, tungod kay kini nagpasiugda sa daghang makadaut nga mga proseso nga mahimong dungan nga ma-target sa usa ka hit sa NRF2, lakip usab ang oxidative stress, neuroinflammation o mitochondrial dysfunction. Bisan pa, gikinahanglan ang umaabot nga trabaho aron siguradong mahibal-an kung ang pagpaaktibo sa pharmacological sa NRF2 mahimo’g usa ka balido nga estratehiya aron mapadali ang pagkadaot sa makahilo nga mga protina sa utok.
Sama sa gipasabut kaniadto, gipasamot ang GSK-3? Ang kalihokan gitaho sa mga sakit nga neurodegenerative ug kini gibanabana nga ang sangputanan nga pagkunhod sa NRF2 mahimong bahin nga responsable sa makadaot nga sangputanan. Ubos niining mga kondisyon sa patolohiya, ang mga inhibitor sa GSK-3 mahimo usab nga magtinabangay aron madugangan ang lebel sa NRF2 ug proteostasis. Ang mapuslanon nga mga epekto sa GSK-3 inhibitors gitaho sa lain-laing mga modelo sa neurodegeneration ug, mas makapaikag, GSK-3 pagpanumpo gipakita sa pagpakunhod sa lebel sa makahilo nga mga protina [130], [131], [132], [133]. Bisan tuod wala'y direktang mga sumpay tali sa GSK-3 nga pagdili ug NRF2-transcriptional nga regulasyon sa mga gene nga nagpasiugda sa proteostasis nga naobserbahan pa, makatarunganon nga pangagpas nga ang down-regulation sa GSK-3 nga kalihokan moresulta sa pagtaas sa lebel sa NRF2, nga sa ngadto-ngadto moresulta sa reinforced. proteostasis.
Ang kalihokan sa transkripsyon sa NRF2 ingon man ang kapasidad sa cellular aron mapadayon ang proteostasis nga pagkunhod sa edad, ang panguna nga hinungdan sa peligro alang sa pag-uswag sa mga sakit nga neurodegenerative. Makatarunganon nga hunahunaon nga ang pagpalig-on sa NRF2 ug, sa ingon, ang proteostasis, labing menos, maglangan sa pagtipon sa mga aggregate sa protina ug neurodegeneration. Sa tinuud, ang pagtambal sa mga senescent fibroblast sa tawo nga adunay 18?-glycyrrhetinic acid (18?-GA) triterpenoid nagpasiugda sa pagpaaktibo sa NRF2, nga nagdala sa proteasome induction ug gipauswag ang gitas-on sa kinabuhi. Kini nga pagtuon nagsugyot nga ang pharmacological activation sa NRF2 posible bisan sa ulahing bahin sa kinabuhi [86]. Dugang pa, ang usa ka ulahi nga pagtuon nagpakita nga kini nga compound nagpataliwala sa SKN-1 ug proteasome nga pagpaaktibo sa C.elegans nga adunay mapuslanon nga mga epekto sa pag-uswag sa AD sa may kalabutan nga mga modelo sa nematode [134].
Ang tanan nga mga butang nga gikonsiderar, ang NRF2-mediated induction sa proteostasis-related nga mga gene daw mapuslanon sa lain-laing mga proteinopathies.
Sulforaphane ug ang mga Epekto Niini sa Kanser, Pagka-mortal, Pagkatigulang, Utok ug Panggawi, Sakit sa Kasingkasing ug uban pa
Ang Isothiocyanates mao ang pipila sa labing hinungdanon nga mga compound sa tanum nga makuha nimo sa imong pagkaon. Niini nga video gihimo nako ang labing komprehensibo nga kaso alang kanila nga nahimo sukad. Mubo nga gidugayon sa pagtagad? Laktaw sa imong paborito nga hilisgutan pinaagi sa pag-klik sa usa sa mga punto sa oras sa ubos. Full timeline sa ubos.
Pangunang mga seksyon:
00:01:14 – Kanser ug mortalidad
00:19:04 - Pagkatigulang
00:26:30 - Utok ug pamatasan
00:38:06 - Katapusan nga recap
00:40:27 - Dosis
Tibuok timeline:
00:00:34 - Pagpaila sa sulforaphane, usa ka mayor nga focus sa video.
00:01:14 - Pagkonsumo sa cruciferous nga utanon ug pagkunhod sa tanan nga hinungdan sa pagkamatay.
00:02:12 - Risgo sa kanser sa prostate.
00:02:23 - Risgo sa kanser sa pantog.
00:02:34 - Ang kanser sa baga sa mga nanigarilyo peligro.
00:02:48 - Risgo sa kanser sa suso.
00:03:13 - Hypothetical: unsa man kung ikaw adunay kanser? (interbensyonal)
00:03:35 - Katuohan nga mekanismo nga nagmaneho sa kanser ug mortalidad nga asosasyon nga datos.
00:04:38 - Sulforaphane ug kanser.
00:05:32 - Ang ebidensya sa hayop nga nagpakita sa kusog nga epekto sa broccoli sprout extract sa pag-uswag sa tumor sa pantog sa mga ilaga.
00:06:06 - Epekto sa direktang suplemento sa sulforaphane sa mga pasyente sa kanser sa prostate.
00:07:09 - Bioaccumulation sa isothiocyanate metabolites sa aktwal nga tisyu sa dughan.
00:08:32 - Pagpugong sa mga stem cell sa kanser sa suso.
00:08:53 - Leksyon sa kasaysayan: ang mga brassicas gitukod nga adunay mga kabtangan sa kahimsog bisan sa karaang Roma.
00:09:16 - Ang abilidad sa Sulforaphane sa pagpalambo sa carcinogen excretion (benzene, acrolein).
00:10:10 - Sa unsang paagi ang pagpaaktibo sa NRF2 makapauswag sa carcinogen excretion pinaagi sa glutathione-S-conjugates.
00:10:34 - Ang Brussels sprouts nagdugang sa glutathione-S-transferase ug makapamenos sa kadaot sa DNA.
00:11:20 - Ang broccoli sprout nga ilimnon nagdugang sa benzene excretion sa 61%.
00:13:31 - Ang homogenate nga pagtubo sa broccoli nagdugang antioxidant enzymes sa taas nga agianan sa hangin.
00:15:45 - Pagkonsumo sa cruciferous nga utanon ug pagkamatay sa sakit sa kasingkasing.
00:16:55 - Ang broccoli sprout powder nagpauswag sa mga lipid sa dugo ug sa kinatibuk-ang risgo sa sakit sa kasingkasing sa type 2 nga mga diabetes.
00:19:04 - Pagsugod sa seksyon sa pagkatigulang.
00:19:21 - Sulforaphane-enriched nga pagkaon nagpauswag sa kinabuhi sa mga bakukang gikan sa 15 ngadto sa 30% (sa pipila ka mga kondisyon).
00:20:34 - Ang kamahinungdanon sa ubos nga panghubag alang sa taas nga kinabuhi.
00:22:05 - Ang cruciferous nga mga utanon ug broccoli sprout powder daw makapakunhod sa nagkalainlain nga mga marker sa panghubag sa mga tawo.
00:23:40 - Mid-video recap: kanser, mga seksyon sa pagkatigulang
00:24:14 - Ang mga pagtuon sa mouse nagsugyot nga ang sulforaphane mahimong makapauswag sa adaptive immune function sa pagkatigulang.
00:25:18 - Ang Sulforaphane nagpauswag sa pagtubo sa buhok sa usa ka modelo sa mouse sa pagkalbo. Hulagway sa 00:26:10.
00:26:30 - Pagsugod sa seksyon sa utok ug pamatasan.
00:27:18 - Epekto sa broccoli sprout extract sa autism.
00:27:48 - Epekto sa glucoraphanin sa schizophrenia.
00:28:17 - Pagsugod sa diskusyon sa depresyon (katuohan nga mekanismo ug pagtuon).
00:31:21 - Ang pagtuon sa mouse gamit ang 10 ka lain-laing mga modelo sa stress-induced depression nagpakita sa sulforaphane nga parehas nga epektibo sama sa fluoxetine (prozac).
00:32:00 - Gipakita sa pagtuon nga ang direkta nga pag-inom sa glucoraphanin sa mga ilaga parehas nga epektibo sa pagpugong sa depresyon gikan sa sosyal nga kapildihan nga modelo sa stress.
00:33:01 - Pagsugod sa seksyon sa neurodegeneration.
00:33:30 - Sulforaphane ug Alzheimer's disease.
00:33:44 - Sulforaphane ug Parkinson's disease.
00:33:51 - Sulforaphane ug sakit sa Hungtington.
00:34:13 - Ang Sulforaphane nagdugang sa mga protina sa heat shock.
00:34:43 - Pagsugod sa seksyon sa traumatic brain injury.
00:35:01 - Sulforaphane gi-injected dayon human ang TBI nagpalambo sa memorya (pagtuon sa mouse).
00:35:55 - Sulforaphane ug neuronal plasticity.
00:36:32 - Ang Sulforaphane nagpauswag sa pagkat-on sa modelo sa type II diabetes sa mga ilaga.
00:37:19 - Sulforaphane ug duchenne muscular dystrophy.
00:37:44 - Pagpugong sa Myostatin sa mga selula sa satellite sa kalamnan (in vitro).
00:38:06 - Late-video recap: mortalidad ug kanser, kadaot sa DNA, oxidative stress ug panghubag, benzene excretion, sakit sa cardiovascular, type II diabetes, mga epekto sa utok (depresyon, autism, schizophrenia, neurodegeneration), NRF2 nga agianan.
00:40:27 - Mga hunahuna sa paghunahuna sa usa ka dosis sa broccoli sprouts o sulforaphane.
00:41:01 - Mga anekdota sa pagtubo sa balay.
00:43:14 - Sa temperatura sa pagluto ug kalihokan sa sulforaphane.
00:43:45 - Ang pagkakabig sa bakterya sa gut sa sulforaphane gikan sa glucoraphanin.
00:44:24 - Ang mga suplemento mas maayo kung gihiusa sa aktibo nga myrosinase gikan sa mga utanon.
00:44:56 - Mga teknik sa pagluto ug cruciferous nga mga utanon.
00:46:06 - Isothiocyanates isip goitrogens.
Ang nuclear factor nga erythroid-derived 2 (NF-E2)-related factor 2, o nailhan nga Nrf2, usa ka transcription factor nga nag-regulate sa ekspresyon sa lain-laing antioxidant ug detoxifying enzymes. Gipakita usab sa mga pagtuon sa panukiduki ang papel niini sa pagkontrol sa stress sa oxidative. Kadaghanan sa mga sakit nga neurodegenerative, sama sa Alzheimer's disease ug Parkinson's disease, gihulagway pinaagi sa oxidative stress ug chronic inflammation, ang kasagarang mga target sa Mga pamaagi sa pagtambal sa Nrf2. Dr. Alex Jimenez DC, CCST Insight
Panapos nga Pulong
Ang transcription factor nga NRF2 nag-orchestrate sa usa ka proteostatic nga tubag pinaagi sa pagbati ug pag-modulate sa mga pagbag-o sa UPR, UPS ug autophagy (Fig. 4). Tungod niini, ang kakulang sa NRF2 gipakita nga nagpasamot sa proteinopathy, nga nagsugyot nga ang NRF2 gikinahanglan alang sa labing maayo nga clearance sa protina. Sa tingub, mahimo natong pangagpas nga ang NRF2 mahimo’g usa ka makapaikag nga target sa pagtambal alang sa mga protinaopathies.
Figure 4 NRF2 isip hub nga nagkonektar sa proteotoxic-derived emergency signal ngadto sa protective transcriptional response. Ang panagtapok sa mga unfolded/misfolded proteins mosangpot sa pagpaaktibo sa unfolded protein response (UPR) sa ER. Ang pagpaaktibo sa PERK o MAPK mahimong moresulta sa transcriptional induction sa ER-resident Gpx8 ug daghang mga enzyme nga nag-regulate sa lebel sa GSH, kritikal aron masiguro ang husto nga pagpilo sa protina. Ang mga aggregate sa protina nagpugong sa kalihokan sa proteasome (UPS), tingali naglikay sa pagkadaot sa NRF2. Ang NRF2 gipakita nga espesipikong modulate sa transkripsyon sa Psma3, Psma6, Psmb1, Psmb5 ug Pomp genes. Daghang uban pang mga subunit ang gi-regulate sa usa ka paagi nga nagsalig sa NRF2 isip tubag sa D3T, tingali gipadak-an ang lista sa mga subunit nga proteasome nga gi-regulate sa NRF2. Ang Autophagy mao ang panguna nga agianan alang sa pagkadaot sa mga aggregate sa protina. Ang Autophagy usab nag-regulate sa NRF2, nga nagkonektar niini nga degradasyon nga agianan sa NRF2 transcriptional induction sa p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 ug Gabarapl1.
Sumala sa artikulo sa ibabaw, samtang ang mga sintomas sa neurodegenerative nga mga sakit mahimong matambalan pinaagi sa lainlaing mga opsyon sa pagtambal, gipakita sa mga pagtuon sa panukiduki nga ang pagpaaktibo sa Nrf2 mahimong usa ka makatabang nga pamaagi sa pagtambal. Kay Ang mga activator sa Nrf2 nagpunting sa halapad nga mga mekanismo sa sakit, ang tanan nga neurodegenerative nga mga sakit makabenepisyo gikan sa paggamit sa Nrf2 transcription factor. Ang mga nahibal-an sa Nrf2 nagbag-o sa pagtambal sa mga sakit nga neurodegenerative. Ang sakup sa among kasayuran limitado sa mga isyu sa kahimsog sa chiropractic ug spinal. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Dugang nga Hisgutan sa Hilisgutan: Paghupay sa Sakit sa Tuhod nga walay Operasyon
Ang sakit sa tuhod usa ka ilado nga simtomas nga mahimong mahitabo tungod sa lain-laing mga samad sa tuhod ug/o kondisyon, lakip nasports injuries. Ang tuhod mao ang usa sa labing komplikado nga mga lutahan sa lawas sa tawo tungod kay kini gilangkoban sa intersection sa upat ka bukog, upat ka ligaments, lain-laing mga tendon, duha ka menisci, ug cartilage. Sumala sa American Academy of Family Physicians, ang kasagarang hinungdan sa kasakit sa tuhod naglakip sa patellar subluxation, patellar tendinitis o jumper's knee, ug Osgood-Schlatter disease. Bisan kung ang sakit sa tuhod lagmit nga mahitabo sa mga tawo nga kapin sa 60 ka tuig ang edad, ang sakit sa tuhod mahimo usab nga mahitabo sa mga bata ug mga tin-edyer. Ang sakit sa tuhod mahimong matambalan sa balay subay sa mga pamaagi sa RICE, bisan pa, ang grabe nga mga samad sa tuhod mahimong magkinahanglan dayon nga medikal nga atensyon, lakip ang pag-atiman sa chiropractic. �
Ang stress sa oxidative gihulagway nga kadaot sa selula nga gipahinabo sa mga libre nga radikal, o dili lig-on nga mga molekula, nga sa katapusan makaapekto sa kahimsog sa kahimsog. Ang lawas sa tawo nagmugna og mga libreng radicals aron sa pag-neutralize sa mga bakterya ug mga virus, bisan pa, ang mga eksternal nga mga hinungdan, sama sa oxygen, polusyon, ug radiation, sa kasagaran makahimo usab og mga libreng radical. Ang tensiyon sa oxidative nalangkit sa daghang mga isyu sa panglawas.
Ang oxidative stress ug uban pang mga stressor nagpalihok sa internal nga mga mekanismo sa pagpanalipod nga makatabang sa pag-regulate sa tubag sa antioxidant sa lawas sa tawo. Ang Nrf2 usa ka protina nga makamatikod sa lebel sa oxidative stress ug makapahimo sa mga selula sa pagpanalipod sa ilang kaugalingon gikan sa internal ug external nga mga hinungdan. Gipakita usab ang Nrf2 nga makatabang sa pag-regulate sa mga gene nga nalambigit sa paghimo sa mga antioxidant nga enzyme ug mga gene nga tubag sa stress. Ang katuyoan sa artikulo sa ubos mao ang pagpatin-aw sa mga epekto sa Nrf2 sa kanser.
abstract
Ang Keap1-Nrf2 nga agianan mao ang nag-unang regulator sa cytoprotective nga mga tubag sa oxidative ug electrophilic stress. Bisan kung ang mga agianan sa pagsenyas sa cell nga gipahinabo sa transcription factor nga Nrf2 nagpugong sa pagsugod ug pag-uswag sa kanser sa normal ug premalignant nga mga tisyu, sa hingpit nga malignant nga mga selula Ang kalihokan sa Nrf2 naghatag kaayohan sa pagtubo pinaagi sa pagdugang sa chemoresistance sa kanser ug pagpauswag sa pagtubo sa selula sa tumor. Niini nga graphical nga pagrepaso, naghatag kami usa ka kinatibuk-ang ideya sa Keap1-Nrf2 nga agianan ug ang dysregulation niini sa mga selula sa kanser. Gi-summarize usab namo ang mga sangputanan sa constitutive Nrf2 nga pagpaaktibo sa mga selula sa kanser ug kung giunsa kini mapahimuslan sa therapy sa gene sa kanser.
keywords:Nrf2, Keap1, Kanser, Antioxidant nga tubag nga elemento, Gene therapy
Pasiuna
Ang Keap1-Nrf2 nga agianan mao ang mayor nga regulator sa cytoprotective nga mga tubag sa endogenous ug exogenous nga mga stress nga gipahinabo sa reactive oxygen species (ROS) ug electrophiles [1]. Ang yawe nga signaling proteins sulod sa agianan mao ang transcription factor Nrf2 (nuclear factor erythroid 2-related factor 2) nga nagbugkos uban sa gagmay nga mga protina sa Maf ngadto sa antioxidant response element (ARE) sa regulatory regions sa target genes, ug Keap1 (Kelch ECH). nakig-uban sa protina 1), usa ka repressor nga protina nga nagbugkos sa Nrf2 ug nagpasiugda sa pagkadaut niini pinaagi sa ubiquitin proteasome nga agianan (Fig. 1). Ang Keap1 usa ka protina nga puno sa cysteine , ang mouse Keap1 adunay kinatibuk-an nga 25 ug 27 nga residu sa cysteine sa tawo, kadaghanan niini mahimong mabag-o sa vitro sa lainlaing mga oxidant ug electrophile [2]. Tulo niini nga mga residues, C151, C273 ug C288, gipakita nga nagdula sa usa ka functional nga papel pinaagi sa pag-usab sa conformation sa Keap1 paingon sa nukleyar nga translocation sa Nrf2 ug sa sunod nga target nga gene expression [3] (Fig. 1). Ang eksakto nga mekanismo diin ang mga pagbag-o sa cysteine sa Keap1 mosangpot sa pagpaaktibo sa Nrf2 wala mahibal-an, apan ang duha nga nagpatigbabaw apan dili managsama nga eksklusibo nga mga modelo mao ang (1) ang �hinge ug latch� nga modelo, diin ang mga pagbag-o sa Keap1 sa mga residu sa thiol nga nagpuyo sa IVR sa Keap1 makabalda sa interaksyon sa Nrf2 hinungdan sa usa ka misalignment sa lysine residues sulod sa Nrf2 nga dili na mahimong polyubiquitinylated ug (2) ang modelo diin ang thiol modification hinungdan sa dissociation sa Cul3 gikan sa Keap1 [3]. Sa duha nga mga modelo, ang inducer-modified ug Nrf2-bound Keap1 dili aktibo ug, tungod niini, ang bag-ong synthesized nga Nrf2 nga mga protina nag-bypass sa Keap1 ug gibalhin ngadto sa nucleus, nagbugkos sa ARE ug nagduso sa ekspresyon sa Nrf2 target nga mga gene sama sa NAD (P) H quinone oxidoreductase 1 (NQO1), heme oxygenase 1 (HMOX1), glutamate-cysteine ligase (GCL) ug glutathione S transferases (GSTs) (Fig. 2). Dugang pa sa mga pagbag-o sa Keap1 thiols nga miresulta sa Nrf2 target gene induction, ang mga protina sama sa p21 ug p62 mahimong makagapos sa Nrf2 o Keap1 sa ingon makabalda sa interaksyon tali sa Nrf2 ug Keap1 [1], [3] (Fig. 3).
Fig. 1. Mga istruktura sa Nrf2 ug Keap1 ug ang cysteine code. (A) Ang Nrf2 naglangkob sa 589 amino acid ug adunay unom ka ebolusyonaryong gikonserbar nga mga domain, Neh1-6. Ang Neh1 adunay usa ka bZip motif, usa ka sukaranan nga rehiyon � leucine zipper (L-Zip) nga istruktura, diin ang sukaranan nga rehiyon mao ang responsable sa pag-ila sa DNA ug ang L-Zip nagpataliwala sa dimerisasyon sa gagmay nga mga protina sa Maf. Ang Neh6 naglihok isip usa ka degron aron sa pagpataliwala sa pagkadaut sa Nrf2 sa nucleus. Ang Neh4 ug 5 maoy mga domain sa transactivation. Ang Neh2 naglangkob sa ETGE ug DLG nga mga motif, nga gikinahanglan alang sa interaksyon sa Keap1, ug usa ka hydrophilic nga rehiyon sa lysine residues (7 K), nga gikinahanglan alang sa Keap1-dependent polyubiquitination ug degradation sa Nrf2. (B) Keap1 naglangkob sa 624 amino acid residues ug adunay lima ka mga domain. Ang duha ka mga motibo sa interaksyon sa protina, ang BTB domain ug ang Kelch domain, gibulag sa intervening region (IVR). Ang domain sa BTB kauban ang N-terminal nga bahin sa IVR nagpataliwala sa homodimerization sa Keap1 ug nagbugkos sa Cullin3 (Cul3). Ang Kelch domain ug ang C-terminal nga rehiyon nagpataliwala sa interaksyon sa Neh2. (C) Nrf2 nakig-uban sa duha ka molekula sa Keap1 pinaagi sa iyang Neh2 ETGE ug DLG motifs. Parehong ETGE ug DLG nagbugkos sa parehas nga mga site sa ilawom nga bahin sa Keap1 Kelch motif. (D) Ang Keap1 dato sa mga residu sa cysteine, nga adunay 27 ka cysteine sa protina sa tawo. Ang uban niini nga mga cysteine nahimutang duol sa mga nag-unang residues ug busa maayo kaayo nga mga target sa mga electrophile ug mga oxidant. Ang sumbanan sa pagbag-o sa mga residu sa cysteine pinaagi sa mga electrophile nailhan nga cysteine code. Ang hypothesis sa cysteine code nagsugyot nga ang lainlaing istruktura nga mga ahente sa pagpaaktibo sa Nrf2 makaapekto sa lainlaing mga cysteine sa Keap1. Ang mga pagbag-o sa cysteine nga nagdala sa mga pagbag-o sa conformational sa Keap1 nga nakabalda sa interaksyon tali sa Nrf2 DLG ug Keap1 Kelch nga mga domain, sa ingon nagpugong sa polyubiquitination sa Nrf2. Ang functional importansya sa Cys151, Cys273 ug Cys288 gipakita, tungod kay gikinahanglan ang Cys273 ug Cys288 alang sa pagsumpo sa Nrf2 ug Cys151 alang sa pagpaaktibo sa Nrf2 sa mga inducers [1], [3].
Fig. 2. Ang Nrf2-Keap1 signaling pathway. (A ug B) sa basal nga mga kondisyon, duha ka Keap1 nga mga molekula nagbugkos sa Nrf2 ug Nrf2 kay polyubiquitylated sa Cul3-based E3 ligase complex. Kini nga polyubiquitilation moresulta sa paspas nga Nrf2 degradation sa proteasome. Ang usa ka gamay nga bahin sa Nrf2 nakalingkawas sa inhibitory complex ug natipon sa nucleus aron sa pagpataliwala sa basal nga ARE-dependent nga ekspresyon sa gene, sa ingon nagmintinar sa cellular homeostasis. (C) Ubos sa mga kondisyon sa tensiyon, ang mga inducers nag-usab sa Keap1 cysteinees nga nagdala ngadto sa pagdili sa Nrf2 ubiquitylation pinaagi sa dissociation sa inhibitory complex. (D) Sumala sa hinge ug latch nga modelo, ang pagbag-o sa piho nga Keap1 cysteine residues mosangpot sa conformational nga mga kausaban sa Keap1 nga miresulta sa detatsment sa Nrf2 DLG motif gikan sa Keap1. Ang Ubiquitination sa Nrf2 nabalda apan ang pagbugkos sa ETGE motif nagpabilin. (E) Sa Keap1-Cul3 dissociation model, ang pagbugkos sa Keap1 ug Cul3 nabalda agig tubag sa mga electrophile, nga mitultol sa pag-ikyas sa Nrf2 gikan sa ubiquitination system. Sa duha sa gisugyot nga mga modelo, ang inducer-modified ug Nrf2-bound Keap1 dili aktibo ug, sa ingon, ang bag-ong synthesized nga Nrf2 nga mga protina nag-bypass sa Keap1 ug gibalhin ngadto sa nucleus, nagbugkos sa Antioxidant Response Element (ARE) ug nagduso sa ekspresyon sa Nrf2 nga target. mga gene sama sa NQO1, HMOX1, GCL ug GSTs [1], [3].
Fig. 3. Mga mekanismo alang sa constitutive nuclear accumulation sa Nrf2 sa cancer. (A) Somatic mutations sa Nrf2 o Keap1 makabalda sa interaksyon niining duha ka protina. Sa Nrf2, ang mga mutation makaapekto sa ETGE ug DLG motifs, apan sa Keap1 mutations mas parehas nga giapod-apod. Dugang pa, ang pagpaaktibo sa oncogene, sama sa KrasG12D [5], o pagkabalda sa mga suppressor sa tumor, sama sa PTEN [11] mahimong mosangpot sa transcriptional induction sa Nrf2 ug pagtaas sa nukleyar nga Nrf2. (B) Ang hypermethylation sa Keap1 promoter sa kanser sa baga ug prostate mosangpot sa pagkunhod sa Keap1 mRNA nga ekspresyon, nga nagdugang sa nukleyar nga akumulasyon sa Nrf2 [6], [7]. (C) Sa familial papillary renal carcinoma, ang pagkawala sa fumarate hydratase enzyme nga kalihokan modala ngadto sa pagtipon sa fumarate ug dugang sa succination sa Keap1 cysteine residues (2SC). Kini nga post-translational nga pagbag-o nagdala ngadto sa pagkabalda sa Keap1-Nrf2 nga interaksyon ug nukleyar nga akumulasyon sa Nrf2 [8], [9]. (D) Ang pagtipon sa mga protina sa disruptor sama sa p62 ug p21 mahimong makadisturbo sa Nrf2-Keap1 nga pagbugkos ug moresulta sa pagtaas sa nukleyar nga Nrf2. Ang p62 nagbugkos sa Keap1 nga nagsapaw sa nagbugkos nga bulsa alang sa Nrf2 ug ang p21 direkta nga nakig-uban sa DLG ug ETGE nga mga motif sa Nrf2, sa ingon nakigkompetensya sa Keap1 [10].
Mga Mekanismo sa Pagpaaktibo ug Dysregulation sa Nrf2 sa Kanser
Bisan kung ang cytoprotection nga gihatag sa pagpaaktibo sa Nrf2 hinungdanon alang sa chemoprevention sa kanser sa normal ug premalignant nga mga tisyu, sa bug-os nga malignant nga mga selula Ang kalihokan sa Nrf2 naghatag kaayohan sa pagtubo pinaagi sa pagdugang sa chemoresistance sa kanser ug pagpauswag sa pagtubo sa selula sa tumor [4]. Daghang mga mekanismo diin ang Nrf2 signaling pathway constitutively activated sa nagkalain-laing mga kanser gihulagway: (1) somatic mutations sa Keap1 o ang Keap1 binding domain sa Nrf2 nga nakabalda sa ilang interaksyon; (2) epigenetic silencing sa Keap1 nga ekspresyon nga nagdala ngadto sa depekto nga pagsumpo sa Nrf2; (3) pagtipon sa mga protina sa disruptor sama sa p62 nga nagdala ngadto sa dissociation sa Keap1-Nrf2 complex; (4) transcriptional induction sa Nrf2 pinaagi sa oncogenic K-Ras, B-Raf ug c-Myc; ug (5) post-translational modification sa Keap1 cysteinees pinaagi sa succinylation nga mahitabo sa familial papillary renal carcinoma tungod sa pagkawala sa fumarate hydratase enzyme activity [3], [4], [5], [6], [7], [ 8], [9], [10] (Fig. 3). Ang constitutively abundant nga protina sa Nrf2 maoy hinungdan sa dugang nga pagpahayag sa mga gene nga nalambigit sa metabolismo sa droga sa ingon nagdugang ang resistensya sa chemotherapeutic nga mga tambal ug radiotherapy. Dugang pa, ang taas nga lebel sa protina sa Nrf2 nalangkit sa dili maayo nga prognosis sa kanser [4]. Ang sobrang aktibo nga Nrf2 makaapekto usab sa pagdaghan sa selula pinaagi sa pagdirekta sa glucose ug glutamine ngadto sa mga anabolic pathway nga nagpadako sa purine synthesis ug nag-impluwensya sa pentose phosphate nga agianan aron sa pagpalambo sa cell proliferation [11] (Fig. 4).
Fig. 4. Ang doble nga papel sa Nrf2 sa tumorigenesis. Ubos sa mga kondisyon sa pisyolohikal, ang ubos nga lebel sa nukleyar nga Nrf2 igo na alang sa pagpadayon sa cellular homeostasis. Gipugngan sa Nrf2 ang pagsugod sa tumor ug metastasis sa kanser pinaagi sa pagwagtang sa mga carcinogens, ROS ug uban pang mga ahente nga makadaot sa DNA. Atol sa tumorigenesis, ang pagtipon sa kadaot sa DNA mosangpot sa constitutive hyperactivity sa Nrf2 nga makatabang sa mga autonomous malignant nga mga selula nga makaagwanta sa taas nga lebel sa endogenous ROS ug aron malikayan ang apoptosis. Ang padayon nga taas nga lebel sa nukleyar nga Nrf2 nagpalihok sa mga metabolikong gene dugang sa mga cytoprotective nga mga gene nga nakatampo sa metabolic reprogramming ug gipauswag nga pagdaghan sa selula. Ang mga kanser nga adunay taas nga lebel sa Nrf2 nalangkit sa dili maayo nga prognosis tungod sa radyo ug chemoresistance ug agresibo nga pagdaghan sa selula sa kanser. Busa, ang kalihokan sa Nrf2 nga agianan nagpanalipod sa unang mga hugna sa tumorigenesis, apan makadaot sa ulahing mga yugto. Busa, alang sa paglikay sa kanser, ang pagpauswag sa kalihokan sa Nrf2 nagpabilin nga usa ka importante nga pamaagi samtang alang sa pagtambal sa kanser, ang Nrf2 pagdili mao ang tilinguhaon [4], [11].
Tungod kay ang taas nga kalihokan sa Nrf2 kasagarang mahitabo sa mga selula sa kanser nga adunay dili maayo nga mga resulta, adunay panginahanglan alang sa mga terapiya nga makapugong sa Nrf2. Ikasubo, tungod sa pagkaparehas sa istruktura sa ubang mga miyembro sa pamilya sa bZip, ang pag-uswag sa piho nga mga inhibitor sa Nrf2 usa ka mahagiton nga buluhaton ug pipila ra nga mga pagtuon sa pagdili sa Nrf2 ang gipatik hangtod karon. Pinaagi sa pag-screen sa natural nga mga produkto, Ren et al. [12] nagpaila sa usa ka antineoplastic compound brusatol isip usa ka Nrf2 inhibitor nga nagpalambo sa chemotherapeutic efficacy sa cisplatin. Dugang pa, ang PI3K inhibitors [11], [13] ug Nrf2 siRNA [14] gigamit sa pagpugong sa Nrf2 sa mga selula sa kanser. Bag-ohay lang, migamit kami og alternatibong pamaagi, nailhan nga cancer suicide gene therapy, aron ma-target ang mga selula sa kanser nga adunay taas nga lebel sa Nrf2. Ang Nrf2-driven lentiviral vectors [15] nga adunay thymidine kinase (TK) gibalhin ngadto sa mga selula sa kanser nga adunay taas nga kalihokan sa ARE ug ang mga selula gitambalan sa usa ka pro-drug, ganciclovir (GCV). Ang GCV gi-metabolize ngadto sa GCV-monophosphate, nga dugang nga phosphorylated sa cellular kinases ngadto sa makahilo nga triphosphate nga porma [16] (Fig. 5). Nagdala kini sa epektibo nga pagpatay dili lamang sa TK nga adunay sulud nga mga selula sa tumor, apan usab sa mga silingan nga mga selyula tungod sa epekto sa pagtan-aw [17]. Ang ARE-regulated TK / GCV gene therapy mahimo nga dugang nga mapauswag pinaagi sa paghiusa sa usa ka cancer chemotherapeutic agent doxorubicin sa pagtambal [16], pagsuporta sa ideya nga kini nga pamaagi mahimong mapuslanon kauban ang tradisyonal nga mga terapiya.
Fig. 5. Suicide gene therapy. Ang constitutive Nrf2 nukleyar nga akumulasyon sa mga selula sa kanser mahimong mapahimuslan pinaagi sa paggamit sa Nrf2-driven viral vector alang sa cancer suicide gene therapy [16]. Niini nga pamaagi, ang lentiviral vector (LV) nga nagpahayag sa thymidine kinase (TK) ubos sa dyutay nga SV40 nga tigpasiugda nga adunay upat ka ARE gibalhin ngadto sa mga selula sa adenocarcinoma sa baga. Ang taas nga lebel sa nukleyar nga Nrf2 nagdala sa lig-on nga pagpahayag sa TK pinaagi sa pagbugkos sa Nrf2. Ang mga selula dayon gitambalan sa usa ka pro-drug, ganciclovir (GCV), nga gi-phosphorylated sa TK. Ang Triphosphorylated GCV makabalda sa DNA synthesis ug mosangpot sa epektibong pagpatay dili lamang sa TK nga adunay sulod nga tumor cells, kondili sa silingang mga selula usab tungod sa epekto sa nagtan-aw.
Ang Nrf2 usa ka master regulator nga nagpalihok sa paghimo sa kusgan nga mga antioxidant sa lawas sa tawo nga makatabang sa pagwagtang sa stress sa oxidative. Ang nagkalainlain nga antioxidant enzymes, sama sa superoxide dismutase, o SOD, glutathione, ug catalase, gi-activate usab pinaagi sa Nrf2 pathway. Dugang pa, ang pipila ka mga phytochemical sama sa turmeric, ashwagandha, bacopa, green tea, ug milk thistle, nag-activate sa Nrf2. Nakaplagan kana sa mga pagtuon sa panukiduki Pag-aktibo sa Nrf2 natural nga makapauswag sa proteksyon sa cellular ug makapabalik sa balanse sa lawas sa tawo.
Dr. Alex Jimenez DC, CCST Insight
Sulforaphane ug ang mga Epekto Niini sa Kanser, Pagka-mortal, Pagkatigulang, Utok ug Panggawi, Sakit sa Kasingkasing ug uban pa
Ang Isothiocyanates mao ang pipila sa labing hinungdanon nga mga compound sa tanum nga makuha nimo sa imong pagkaon. Niini nga video gihimo nako ang labing komprehensibo nga kaso alang kanila nga nahimo sukad. Mubo nga gidugayon sa pagtagad? Laktaw sa imong paborito nga hilisgutan pinaagi sa pag-klik sa usa sa mga punto sa oras sa ubos. Full timeline sa ubos.
Pangunang mga seksyon:
00:01:14 – Kanser ug mortalidad
00:19:04 - Pagkatigulang
00:26:30 - Utok ug pamatasan
00:38:06 - Katapusan nga recap
00:40:27 - Dosis
Tibuok timeline:
00:00:34 - Pagpaila sa sulforaphane, usa ka mayor nga focus sa video.
00:01:14 - Pagkonsumo sa cruciferous nga utanon ug pagkunhod sa tanan nga hinungdan sa pagkamatay.
00:02:12 - Risgo sa kanser sa prostate.
00:02:23 - Risgo sa kanser sa pantog.
00:02:34 - Ang kanser sa baga sa mga nanigarilyo peligro.
00:02:48 - Risgo sa kanser sa suso.
00:03:13 - Hypothetical: unsa man kung ikaw adunay kanser? (interbensyonal)
00:03:35 - Katuohan nga mekanismo nga nagmaneho sa kanser ug mortalidad nga asosasyon nga datos.
00:04:38 - Sulforaphane ug kanser.
00:05:32 - Ang ebidensya sa hayop nga nagpakita sa kusog nga epekto sa broccoli sprout extract sa pag-uswag sa tumor sa pantog sa mga ilaga.
00:06:06 - Epekto sa direktang suplemento sa sulforaphane sa mga pasyente sa kanser sa prostate.
00:07:09 - Bioaccumulation sa isothiocyanate metabolites sa aktwal nga tisyu sa dughan.
00:08:32 - Pagpugong sa mga stem cell sa kanser sa suso.
00:08:53 - Leksyon sa kasaysayan: ang mga brassicas gitukod nga adunay mga kabtangan sa kahimsog bisan sa karaang Roma.
00:09:16 - Ang abilidad sa Sulforaphane sa pagpalambo sa carcinogen excretion (benzene, acrolein).
00:10:10 - Sa unsang paagi ang pagpaaktibo sa NRF2 makapauswag sa carcinogen excretion pinaagi sa glutathione-S-conjugates.
00:10:34 - Ang Brussels sprouts nagdugang sa glutathione-S-transferase ug makapamenos sa kadaot sa DNA.
00:11:20 - Ang broccoli sprout nga ilimnon nagdugang sa benzene excretion sa 61%.
00:13:31 - Ang homogenate nga pagtubo sa broccoli nagdugang antioxidant enzymes sa taas nga agianan sa hangin.
00:15:45 - Pagkonsumo sa cruciferous nga utanon ug pagkamatay sa sakit sa kasingkasing.
00:16:55 - Ang broccoli sprout powder nagpauswag sa mga lipid sa dugo ug sa kinatibuk-ang risgo sa sakit sa kasingkasing sa type 2 nga mga diabetes.
00:19:04 - Pagsugod sa seksyon sa pagkatigulang.
00:19:21 - Sulforaphane-enriched nga pagkaon nagpauswag sa kinabuhi sa mga bakukang gikan sa 15 ngadto sa 30% (sa pipila ka mga kondisyon).
00:20:34 - Ang kamahinungdanon sa ubos nga panghubag alang sa taas nga kinabuhi.
00:22:05 - Ang cruciferous nga mga utanon ug broccoli sprout powder daw makapakunhod sa nagkalainlain nga mga marker sa panghubag sa mga tawo.
00:23:40 - Mid-video recap: kanser, mga seksyon sa pagkatigulang
00:24:14 - Ang mga pagtuon sa mouse nagsugyot nga ang sulforaphane mahimong makapauswag sa adaptive immune function sa pagkatigulang.
00:25:18 - Ang Sulforaphane nagpauswag sa pagtubo sa buhok sa usa ka modelo sa mouse sa pagkalbo. Hulagway sa 00:26:10.
00:26:30 - Pagsugod sa seksyon sa utok ug pamatasan.
00:27:18 - Epekto sa broccoli sprout extract sa autism.
00:27:48 - Epekto sa glucoraphanin sa schizophrenia.
00:28:17 - Pagsugod sa diskusyon sa depresyon (katuohan nga mekanismo ug pagtuon).
00:31:21 - Ang pagtuon sa mouse gamit ang 10 ka lain-laing mga modelo sa stress-induced depression nagpakita sa sulforaphane nga parehas nga epektibo sama sa fluoxetine (prozac).
00:32:00 - Gipakita sa pagtuon nga ang direkta nga pag-inom sa glucoraphanin sa mga ilaga parehas nga epektibo sa pagpugong sa depresyon gikan sa sosyal nga kapildihan nga modelo sa stress.
00:33:01 - Pagsugod sa seksyon sa neurodegeneration.
00:33:30 - Sulforaphane ug Alzheimer's disease.
00:33:44 - Sulforaphane ug Parkinson's disease.
00:33:51 - Sulforaphane ug sakit sa Hungtington.
00:34:13 - Ang Sulforaphane nagdugang sa mga protina sa heat shock.
00:34:43 - Pagsugod sa seksyon sa traumatic brain injury.
00:35:01 - Sulforaphane gi-injected dayon human ang TBI nagpalambo sa memorya (pagtuon sa mouse).
00:35:55 - Sulforaphane ug neuronal plasticity.
00:36:32 - Ang Sulforaphane nagpauswag sa pagkat-on sa modelo sa type II diabetes sa mga ilaga.
00:37:19 - Sulforaphane ug duchenne muscular dystrophy.
00:37:44 - Pagpugong sa Myostatin sa mga selula sa satellite sa kalamnan (in vitro).
00:38:06 - Late-video recap: mortalidad ug kanser, kadaot sa DNA, oxidative stress ug panghubag, benzene excretion, sakit sa cardiovascular, type II diabetes, mga epekto sa utok (depresyon, autism, schizophrenia, neurodegeneration), NRF2 nga agianan.
00:40:27 - Mga hunahuna sa paghunahuna sa usa ka dosis sa broccoli sprouts o sulforaphane.
00:41:01 - Mga anekdota sa pagtubo sa balay.
00:43:14 - Sa temperatura sa pagluto ug kalihokan sa sulforaphane.
00:43:45 - Ang pagkakabig sa bakterya sa gut sa sulforaphane gikan sa glucoraphanin.
00:44:24 - Ang mga suplemento mas maayo kung gihiusa sa aktibo nga myrosinase gikan sa mga utanon.
00:44:56 - Mga teknik sa pagluto ug cruciferous nga mga utanon.
00:46:06 - Isothiocyanates isip goitrogens.
mga sakripisyo
Kini nga buhat gisuportahan sa Academy of Finland, ang Sigrid Juselius Foundation ug ang Finnish Cancer Organizations.
Sa konklusyon, ang nukleyar nga factor (erythroid-derived 2) -sama sa 2, nailhan usab nga NFE2L2 o Nrf2, usa ka protina nga nagdugang sa produksiyon sa mga antioxidant nga nanalipod sa lawas sa tawo batok sa oxidative stress. Sama sa gihulagway sa ibabaw, ang stimulation sa Nrf2 nga agianan gitun-an alang sa pagtambal sa mga sakit nga gipahinabo sa oxidative stress, lakip ang cancer. Ang sakup sa among kasayuran limitado sa mga isyu sa kahimsog sa chiropractic ug spinal. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Dugang nga Hisgutan sa Hilisgutan: Paghupay sa Sakit sa Tuhod nga walay Operasyon
Ang sakit sa tuhod usa ka ilado nga simtomas nga mahimong mahitabo tungod sa lain-laing mga samad sa tuhod ug/o kondisyon, lakip nasports injuries. Ang tuhod mao ang usa sa labing komplikado nga mga lutahan sa lawas sa tawo tungod kay kini gilangkoban sa intersection sa upat ka bukog, upat ka ligaments, lain-laing mga tendon, duha ka menisci, ug cartilage. Sumala sa American Academy of Family Physicians, ang kasagarang hinungdan sa sakit sa tuhod naglakip sa patellar subluxation, patellar tendinitis o jumper's knee, ug Osgood-Schlatter disease. Bisan kung ang sakit sa tuhod lagmit nga mahitabo sa mga tawo nga kapin sa 60 ka tuig ang edad, ang sakit sa tuhod mahimo usab nga mahitabo sa mga bata ug mga tin-edyer. Ang sakit sa tuhod mahimong matambalan sa balay subay sa mga pamaagi sa RICE, bisan pa, ang grabe nga mga samad sa tuhod mahimong magkinahanglan dayon nga medikal nga atensyon, lakip ang pag-atiman sa chiropractic.
Gisuportahan sa DNA ang gibana-bana nga 20,000 nga mga gene, ang matag usa adunay usa ka programa alang sa paghimo sa usa ka protina o enzyme nga gikinahanglan alang sa usa ka himsog nga estilo sa kinabuhi. Ang matag usa niini nga mga sumbanan kinahanglan nga kanunay nga kontrolahon sa usa ka matang sa "promoter" nga nagdumala sa eksakto kung unsa kadaghan sa matag substansiya ug/o kemikal ang namugna ug kung asa nga mga kondisyon kini usab molambo.
Pinaagi sa pagkonektar sa usa ka partikular nga klase sa switch-like promoter nga mga lugar, nailhan nga Antioxidant Response Element, o ARE, ang Nrf2 nga hinungdan�nagsuporta sa katulin sa paglalang alang sa gatusan ka lahi nga mga gene nga makapaarang sa mga selyula nga mabuhi sa ilawom sa tensiyonado nga mga kahimtang. Kini nga mga gene nagmugna og usa ka pagpili sa antioxidant enzymes nga nagpalambo sa usa ka network sa depensa pinaagi sa pag-neutralize sa mga oxidant ug pinaagi sa paglimpyo sa makahilo nga by-products nga nahabilin sa ilang produksyon, dugang sa pagtabang sa pagpasig-uli sa kadaot nga ilang gipahinabo.
Unsa ang Stress Oxidative?
Daghang mga oxidant sama sa superoxide radical, o O2-., ug hydrogen peroxide, o H2O2, namugna pinaagi sa praktis sa pagsunog sa mga substansiya ug/o mga kemikal nga nagsustento sa lawas sa tawo. Ang lawas sa tawo adunay mga antioxidant nga enzyme nga nag-neutralize ug nag-detoxize sa mga reaktibo nga pagkaon ug ilimnon nga atong giinom. Gibag-o sa Nrf2 ang ilang produksiyon aron mapadayon ang balanse ug gipasiugda ang panginahanglan alang sa tanan niini nga mga enzyme. Kini nga balanse mahimong mabalda sa daghang mga hinungdan, lakip ang edad.
Sa atong pagkatigulang, ang lawas sa tawo nagmugna og gamay nga Nrf2 ug kini nga delikado nga balanse mahimong anam-anam nga magsugod ngadto sa oxidative nga bahin, usa ka estado nga gitawag nga oxidative stress. Ang sakit mahimo usab nga hinungdan sa sobra nga produksiyon sa mga oxidant. Ang mga impeksyon, alerdyi, ug mga sakit sa autoimmune mahimo usab nga mag-aghat sa atong mga immune cells nga maghimo mga reaktibo nga oxidant, sama sa O2-. , H2O2, OH ug HOCl, diin ang himsog nga mga selula madaot ug motubag sa panghubag. Ang mga sakit nga nalangkit sa pagkatigulang, lakip ang pag-atake sa kasingkasing, stroke, kanser, ug mga kondisyon sa neurodegenerative sama sa Alzheimer's disease, nagdugang usab sa pag-uswag sa mga oxidant, nagpatunghag stress ug tubag sa panghubag.
Unsa ang Nrf2 Activators?
Ang Nrf2 nga protina, nga gitawag usab nga transcription factor tungod sa paagi nga kini makasuporta ug makakontrol sa mga enzyme ug mga gene, mao ang sekreto nga elemento sa usa ka han-ay sa biochemical reactions sulod sa selula nga motubag sa mga kausaban sa cognitive equilibrium ingon man sa oxidative balance. Ang sensing nga mga elemento niini nga agianan nag-usab ug nagpagawas sa Nrf2, nga nagpalihok niini aron kini mokaylap ngadto sa nucleus sa selula paingon sa DNA. Ang Nrf2 mahimo usab nga i-on o i-off ang mga gene ug mga enzyme nga gisuportahan niini aron mapanalipdan ang selula.
Maayo na lang, usa ka lainlaing mga substansiya nga mga Nrf2 activator ang naugmad pinaagi sa pagkonsumo sa pipila nga mga tanum ug mga extract nga gigamit mga siglo na ang milabay sa tradisyonal nga mga tambal nga Intsik ug Lumad nga Amerikano. Kini nga mga phytochemical ingon nga parehas nga kusgan nga adunay gamay nga epekto, sama sa Nrf2-activating pharmaceutical nga mga produkto nga gigamit karon.
Ang nuclear factor nga erythroid 2-related factor, nga mas nailhan nga Nrf2, usa ka transcription factor nga nanalipod sa cell pinaagi sa pag-regulate sa mga gene, enzymes ug antioxidant nga mga tubag. Ang mga hinungdan sa transkripsyon usa ka klase sa protina nga gilakip sa DNA aron mapauswag ang paghimo sa mga piho nga sangkap ug kemikal, lakip ang glutathione S-transferases, o GST. Ang pagpaaktibo sa Nrf2 nag-aghat sa paghimo sa mga aktibo nga protina nga nagpakita sa usa ka kusgan nga kapasidad sa antioxidant aron matabangan ang pagkunhod sa stress sa oxidative.
Dr. Alex Jimenez DC, CCST Insight
Ang Siyensya sa Luyo sa Nrf2 Pagpaaktibo
Sa higayon nga ang inisyal nga Nrf2-activating dietary supplement gihimo sa 2004, gamay nga impormasyon ang nahibal-an mahitungod sa function sa Nrf2 pathway. Gibana-bana nga 200 nga mga mantalaan sa literatura sa Nrf2, nailhan usab nga nuclear factor-like 2 o NFE2L2, naglungtad ug ang mga tigdukiduki nagsugod pa lamang sa pagdiskobre sa antioxidant nga tubag sa Nrf2 sa mga mammal. Sa 2017, bisan pa, kapin sa 9,300 nga mga pagtuon sa panukiduki sa akademiko bahin sa kini nga "master regulator," ang naimprinta.
Sa tinuud, ang Nrf2 nag-regulate sa daghang mga antioxidant nga enzyme nga wala’y kalabotan sa mga gene, sa baylo, nagtanyag sila proteksyon batok sa lainlaing mga kahimtang nga may kalabotan sa stress nga nasugatan sa mga selyula, organo ug sa katapusan mga organismo, sa ilawom sa himsog ug patolohiya nga kahimtang. Pinasukad niining bag-ong gidaghanon sa impormasyon gikan sa gipatik nga mga pagtuon sa panukiduki sa akademiko, ang mga tigdukiduki mahimo na nga molambo nga mas maayo Nrf2 nga mga suplemento sa pagkaon.
Ingon sa 2007, ang mga pagtuon sa panukiduki nagpakita sa komplikado nga gimbuhaton sa Nrf2 nga agianan. Ang mga activator sa Nrf2 nakit-an nga nagsundog sa mga hinungdan sa lainlaing mga istruktura sa sulod sa lawas sa tawo. Pinaagi sa kini nga mga agianan, ang mga activator sa Nrf2 nasangkapan aron mabati ang pagbag-o nga mga kahimtang sa tibuuk nga selyula aron mapadayon ang balanse ug pagtubag sa nagbag-o nga mga kinahanglanon sa mga gene.
Ngano nga Gamiton ang Nrf2-Activating Supplements?
Samtang ang mga abilidad sa pagpaaktibo sa Nrf2 mikunhod uban ang edad sa mga organismo, ang mga pagbag-o mahimong magsugod nga mahitabo. Gipakita sa mga pagtuon sa panukiduki nga ang pokus sa Nrf2 sa mga selyula mikunhod uban ang edad, nga nagpakita sa dugang nga mga marka sa oxidative stress. Ang lainlaing mga sakit nga may kalabotan sa edad sama sa atherosclerosis ug sakit sa cardiovascular, arthritis, kanser, sobra nga katambok, type 2 diabetes, hypertension, katarata, ug sakit nga Alzheimer ingon man ang mga sakit sa Parkinson mahimong molambo tungod sa kini nga mga pagbag-o. Ang oxidative stress nakit-an sa kini nga mga isyu sa kahimsog.
Pinaagi sa pagpukaw sa kapasidad sa selula aron madugangan ang produksiyon sa Nrf2 activators, Nrf2 nga mga suplemento sa pagkaon makatabang sa pagpabuhi sa kaugalingong abilidad sa lawas sa tawo sa pagsumpo sa mga epekto sa oxidative stress. Ang mga polyunsaturated fatty acid, o PUFA, usa sa labing dali nga ma-oxidized nga mga molekula ug sila labi ka dali nga maapektuhan sa kadaot gikan sa mga libre nga radikal. Ang Thiobarbituric acid, o TBARS, ang produksiyon mahimong madugangan sa edad, nga nagpaila sa gipataas nga stress sa oxidative kauban ang usa ka pagtulo sa Nrf2-regulated nga mga agianan.
Sa biyolohikal nga paagi, ang gene induction kay hinay kaayo nga mekanismo, kasagaran nagkinahanglan ug mga oras aron mabalhin sa usa ka agianan. Ingon usa ka sangputanan, daghang mga enzyme ang adunay kaugalingon nga on/off switch nga mahimong ma-trigger sa mga minuto sa lainlaing mga regulatory enzyme. Ang mga tigdukiduki nakahimo og proprietary nga komposisyon sa Nrf2 activators nga naggamit niini nga kahibalo nga base sa pagpaaktibo. Ang pagpaaktibo sa Nrf2 gilangkuban dili lamang sa Nrf2 transcription factor nga gipagawas gikan sa inhibitor niini ug migrate ngadto sa cell nucleus, apan nagbugkos usab sa piho nga mga sequence sa DNA aron madasig ang cytoprotective gene expression, nga nag-regulate sa dagan sa Nrf2 nga gikuha gikan sa nucleus.
Ang pagsabut sa pamaagi sa pagwagtang ug ang pagpaaktibo sa Nrf2 sa lawas sa tawo nagtugot sa mga tigdukiduki sa pagtukod og mga kombinasyon sa lain-laing mga Nrf2 activators aron matuman ang pagpamalandong sa mga gene pinaagi sa modulasyon niini. Ang kombinasyon sa base sa kahibalo, kauban ang daghang lainlain nga mga pagtuon sa panukiduki nakatabang sa paghimo sa mga activator sa Nrf2 aron magamit ingon mga suplemento sa pagkaon. Ang sakup sa among kasayuran limitado sa mga isyu sa kahimsog sa chiropractic ug spinal. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Gi-curate ni Dr. Alex Jimenez
Dugang nga Hisgutan sa Hilisgutan: Paghupay sa Sakit sa Tuhod nga walay Operasyon
Ang sakit sa tuhod usa ka ilado nga simtomas nga mahimong mahitabo tungod sa lain-laing mga samad sa tuhod ug/o kondisyon, lakip nasports injuries. Ang tuhod mao ang usa sa labing komplikado nga mga lutahan sa lawas sa tawo tungod kay kini gilangkoban sa intersection sa upat ka bukog, upat ka ligaments, lain-laing mga tendon, duha ka menisci, ug cartilage. Sumala sa American Academy of Family Physicians, ang kasagarang hinungdan sa sakit sa tuhod naglakip sa patellar subluxation, patellar tendinitis o jumper's knee, ug Osgood-Schlatter disease. Bisan kung ang sakit sa tuhod lagmit nga mahitabo sa mga tawo nga kapin sa 60 ka tuig ang edad, ang sakit sa tuhod mahimo usab nga mahitabo sa mga bata ug mga tin-edyer. Ang sakit sa tuhod mahimong matambalan sa balay subay sa mga pamaagi sa RICE, bisan pa, ang grabe nga mga samad sa tuhod mahimong magkinahanglan dayon nga medikal nga atensyon, lakip ang pag-atiman sa chiropractic.
Ang mga antioxidant kay siyentipikanhon nga gitawag nga mga compound nga nagpugong sa proseso sa oksihenasyon sa lawas sa tawo, nga kung dili makontrol, mahimo’g makamugna kini nga mga libre nga radikal nga mahimo’g makamugna og daghang mga kadena nga reaksyon nga mahimong hinungdan sa kadaot sa cellular. Maayo na lang, ang lawas sa tawo makahimo sa ingon nga built-in nga mga mekanismo sa imyunidad, bisan pa, kung ang pag-mount sa mga reactive oxygen species, o ROS, dili ma-neutralize, hunahunaa ang usa ka gamay nga siga nga dili makontrol kung gisudlan sa oxygen, ang kadaot mahitabo. .
Aron mapadayon ang pagpalapad sa metapora sa siga, ang katapusan nga produkto nga wala’y katakus sa pag-neutralize sa epekto sa ROS, o reaktibo nga mga species sa oxygen, mao ang kadaot ingon man panghubag, sa laing pagkasulti, ang lawas sa tawo literal nga nagdilaab. Ang katingad-an nga butang mao ang adunay mga antioxidant nga makatabang kaayo sa pagbatok sa kini nga isyu sa kahimsog ug kini nga antioxidant mao ang glutathione. Bisan kung nakit-an kaniadtong 1889, ang epekto sa antioxidant sa glutathione nahimo nga usa sa labing makapaikag nga mga hilisgutan sa modernong mga pagtuon sa panukiduki.
Agalon sa Antioxidants: Glutathione
Ang kusgan nga substansiya usa ka tripeptide nga naugmad gikan sa cysteine, glutamic acid, ug glycine. Tungod sa katakus niini sa pagpanalipod sa lawas sa tawo batok sa paghimo sa mga libre nga radikal, ang glutathione sa katapusan makatabang sa pagpauswag sa usa ka himsog nga immune system. Base sa Scientific Reports sa 2015, nahibal-an nga ang kapasidad sa glutathione nga molihok nga synergistically sa peroxiredin ug catalase makatabang sa pagbantay sa mga selyula batok sa hydrogen peroxide. Kini nga synergistic nga pormula naglihok batok sa reactive oxygen species, o ROS. Ang glutathione, peroxidredin ug catalase mga importanteng elemento sa pagpadako sa cellular homeostasis, nga usa ka importanteng proseso sa himsog nga mga selula, tisyu ug organo sa kinatibuk-an.
Dugang pa, ang glutathione nagdugang sa kinatibuk-ang istruktura sa immune system ug naglihok gamit ang hinungdanon nga epekto niini sa mga function sa lymphocyte. Sumala sa Departamento sa Immunochemistry, ang husto nga pagdugang sa lebel sa glutathione sa lawas sa tawo makapauswag pag-ayo sa mga reaksiyon sa imyunidad. Pananglitan, ang duha ka randomized placebo-controlled nga mga pagsulay nagpakita nga ang terapyutik nga pagtambal sa immune-compromised nga mga pasyente nga adunay N-acetyl-cysteine, o NAC, miresulta, sa duha ka mga kaso, sa usa ka igo nga pagtubo sa kadaghanan sa mga proseso sa immunological nga naglakip sa usa ka tibuok nga rejuvenation. sa natural nga killer cell nga kalihokan. Ang N-acetyl-cysteine, o NAC, naggamit sa sulfur gikan sa glutathione ug gikombinar kini sa makahilong mga molekula, nga unya mahimong matunaw sa tubig ug ipagawas sa lawas sa tawo.
Ang glutathione usab adunay kapabilidad sa pagpabuhi sa lipoic acid ingon man sa pag-recycle sa Vitamin C ug E, nga gikinahanglan aron masugdan ang pipila ka proseso sa sistema pinaagi sa pagpadala sa mga electron aron ma-neutralize ang mga free radical. Base sa research study gikan sa PLOS ONE, glutathione nga apektado sa mga pasyente nga adunay diabetes metillus, o T2DM, ug mycobacterium tuberculosis. Kasagaran, ang mga indibidwal nga adunay huyang nga sistema sa imyunidad adunay kalagmitan nga magpakita sa labi nga pagkaladlad sa M. tb, o mycobacterium tuberculosis, sakit o impeksyon. Dugang pa, ang mga indibidwal nga adunay Type 2 diabetes metilllus, o T2DM, duha ngadto sa tulo ka pilo nga mas prone sa TB kaysa mga tawo nga walay T2DM. Gisugyot usab sa pagtuon sa panukiduki nga ang pagpataas sa lebel sa glutathione sa mga macrophage nga nahimulag gikan sa mga pasyente nga adunay T2DM misangpot sa pagpauswag sa pagkontrol sa sakit o impeksyon sa M.Tb. Kini nga mga resulta nagpakita nga ang ubos nga lebel sa glutathione sa mga pasyente nga adunay T2DM nakatampo sa usa ka taas nga kahigayonan sa M. tb nga sakit o impeksyon. Dugang pa, nagdepende sa Dietro Ghezzi sa Brighton ug Sussex Medical School, ang oxidative stress sa katapusan mahimong hinungdan sa dili maayo nga istruktura ug function sa immune system.
Maayo na lang, ang glutathione adunay hinungdanon nga papel sa pagpalig-on ug pagpugong sa resistensya. Pananglitan, ang glutathione kinahanglanon alang sa mga natural ug adaptive nga mga proseso sulod sa immune system, lakip ang T-lymphocyte proliferation, phagocytic activity sa polymorphonuclear neutrophils, ug dendritic cell functions, nga mahimong sukaranan tungod kay kini gilangkoban sa antigen-presenting cells. . Ang cell-meditated immunity naglakip sa mga antigens sa protina nga sa sinugdan nagsugod sa pagkadaot sa mga endocytic vesicle sa macrophage ug dendritic nga mga selula, busa, ang gagmay nga mga peptide gipakita sa ibabaw aron ma-aktibo ang pagdaghan sa mga antigen-specific nga T cells. Dugang pa, ang glutathione makatabang sa paghimo sa mga cytokine, ug kinahanglan nga mapadayon ang produksiyon sa interferon-gamma pinaagi sa mga dendritic cells, nga hinungdanon sa pagpanalipod batok sa intracellular pathogens lakip ang mycobacteria.
Ang N-acetyl-cysteine, o NAC, nga gitumong sa siyentipikanhong paagi ingong pasiuna sa glutathione, usa usab ka gamhanan kaayo nga cellular antioxidant nga gigamit isip free radical scavenger antioxidant. Kasagaran nga giila tungod sa papel niini sa paglikay sa acetaminophen toxicity, NAC, o N-acetyl-cysteine, gipakita nga adunay daghang mga benepisyo sa kahimsog ug kahimsog. Sumala sa Cell Journal, ang NAC nagtabang sa pagsuporta sa usa ka himsog nga tubag sa pagpanghubag ug mahimong positibo nga makaapekto sa termino sa tawo ug sa wala pa mahuman nga trabaho. Ang pagtuon sa panukiduki nakahinapos nga sa mga kababayen-an nga adunay nauna nga pagpanganak ug bacterial vaginosis, 0.6 gramo sa NAC kada adlaw nga gikuha sa binaba uban sa progesterone human sa semana nga 16 sa pagmabdos nanalipod batok sa preterm nga pagbalik sa pagpanganak ug pagpalambo sa neonatal nga resulta. Sa konklusyon, ang positibo nga epekto sa NAC sa pagtukod sa kaunuran nakit-an usab. Pagkahuman sa tulo ka minuto nga padayon nga mga kontraksyon, adunay 15 nga porsyento nga gipaayo nga output, nga nagpakita kung giunsa ang NAC adunay hinungdanon nga papel sa pagpauswag sa pagtukod sa kaunuran ug pagkunhod sa kinatibuk-ang kakapoy sa panahon sa paghago.
Nadiskobrehan usab sa mga tigdukiduki nga ang NAC, o N-acetyl-cysteine, mahimong makabenepisyo niadtong adunay polycystic ovarian syndrome, o PCOS. Ang PCOS, o polycystic ovarian syndrome, usa ka komon nga endocrine glands-related nga sakit nga makaapekto sa gibana-bana nga 5 ngadto sa 10 porsyento sa mga babaye nga nag-edad og reproductive. Sa ingon nga mga pasyente, adunay mas dako nga peligro nga makasinati sa metabolic syndrome, diin ang paggamit sa NAC nakatabang sa pagpasig-uli sa himsog nga lebel sa insulin ug pagkasensitibo.
Ang Panabut ni Dr. Alex Jimenez
Ang Glutathione gitawag nga "master of antioxidants" tungod sa sukaranan nga papel niini sa pagkab-ot ug pagmentinar sa kinatibuk-ang kahimsog ug kahimsog. Samtang ang lawas sa tawo makahimo sa paghimo sa kaugalingon nga glutathione, ang dili maayo nga nutrisyon, polusyon, hilo, sobra nga paggamit sa mga tambal ug / o tambal, stress, trauma, pagkatigulang, sakit ug radiation mahimo’g makapakunhod sa atong natural nga lebel sa glutathione. Kini sa baylo makahimo sa mga indibidwal nga mas daling madaot sa selyula gikan sa oxidative stress, free radicals, impeksyon ug kanser. Ang suplemento sa glutathione mahimong adunay daghang mga benepisyo sa lawas sa tawo. Uban sa alternatibong mga opsyon sa pagtambal, sama sa chiropractic care, ang lebel sa glutathione mahimo na usab nga ma-regulate aron mapalambo ang kaayohan.
Dugang pa, gisugyot sa mga propesyonal sa pag-atiman sa kahimsog nga ipatuman ang paggamit sa suplemento sa glutathione kauban ang uban pang mga kapilian sa pagtambal, sama sa pag-atiman sa chiropractic, aron mapalambo pa ang kinatibuk-ang kahimsog ug kahimsog. Ang mga antioxidant hinungdanon sa pagpadayon sa labing taas nga kaayohan ingon man sa pagpugong sa kadena nga reaksyon sa mga libre nga radikal nga hinungdan sa kadaot o kadaot sa selyula. Gamhanan nga mga antioxidant sama sa glutathione, sama sa nahisgotan na sa ibabaw, sa katapusan makatabang sa pag-regulate sa pag-uswag sa kini nga mga libre nga radikal ug maghatag usa ka labi ka himsog nga tubag sa immune system. Nakaplagan kana sa mga pagtuon sa panukiduki pag-atiman sa chiropractic mahimo usab nga adunay hinungdanon nga papel sa kini nga proseso, natural nga nagpadako sa kalihokan sa mga antioxidant sa lawas sa tawo. Ang pag-atiman sa Chiropractic usa ka luwas ug epektibo nga pamaagi sa pagtambal nga naggamit sa spinal adjustments ug manual manipulations aron matul-id ang spinal misalignments, o subluxations, aron tugotan ang lawas sa tawo nga natural nga makaayo sa kaugalingon nga walay paggamit sa mga droga / tambal ug / o surgical interventions.
Sa katapusan, ang mga antioxidant nagpakita sa ilang biological nga mga kabtangan pinaagi sa daghang mga benepisyo sa kahimsog, nga mahimo’g gikinahanglan karon labi pa sa tanan nga nag-uswag nga pag-atake sa tensiyon, sakit ug polusyon sa atong modernong kalibutan, nga ang tanan nakatampo sa kadaot sa cell ug / o kadaot. . Ang glutathione ug ang nag-una niini, NAC, o N-acetyl-cysteine, nagpadayon sa pagpakita sa ilang kusgan nga kahimtang sa gingharian sa mga antioxidant. Uban sa alternatibong mga opsyon sa pagtambal, sama sa pag-atiman sa chiropractic, ang mga tawo makapahimulos sa tanang benepisyo nga ikatanyag niining gamhanang antioxidant. Ang sakup sa among kasayuran limitado sa chiropractic ingon man sa mga kadaot ug kondisyon sa taludtod. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Gi-curate ni Dr. Alex Jimenez
Dugang nga mga Hilisgutan: Sakit sa Bukobuko
Sakit sa likod mao ang usa sa labing kaylap nga hinungdan sa pagkabaldado ug wala’y trabaho nga mga adlaw sa trabaho sa tibuuk kalibutan. Sa tinuud, ang sakit sa bukobuko gipasangil nga ikaduha nga kasagarang hinungdan sa mga pagbisita sa opisina sa doktor, nga mas daghan kaysa mga impeksyon sa taas nga respiratoryo. Gibana-bana nga 80 porsyento sa populasyon ang makasinati og usa ka matang sa sakit sa likod labing menos kausa sa tibuok nilang kinabuhi. Ang dugokan usa ka komplikado nga istruktura nga gilangkoban sa mga bukog, lutahan, ligaments ug kaunuran, ug uban pang humok nga mga tisyu. Tungod niini, mga kadaot ug/o nagkagrabe nga mga kondisyon, sama sa herniated discs, mahimong mosangpot sa mga sintomas sa sakit sa likod. Ang mga samad sa sports o mga aksidente sa awto sa kasagaran mao ang labing kanunay nga hinungdan sa sakit sa bukobuko, bisan pa, usahay ang pinakasimple nga mga paglihok mahimong adunay sakit nga mga sangputanan. Maayo na lang, ang alternatibong mga opsyon sa pagtambal, sama sa pag-atiman sa chiropractic, makatabang sa pagpagaan sa sakit sa likod pinaagi sa paggamit sa mga pag-adjust sa taludtod ug mga pagmaniobra sa manwal, nga sa katapusan makapauswag sa paghupay sa kasakit.
Ang Chiropractor nga nakabase sa siyensya nga si Dr. Alexander Jimenez nagtan-aw sa oxidative stress, unsa kini, kung giunsa kini makaapekto sa lawas ug sa antioxidant nga depensa aron masulbad ang sitwasyon.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 ug Omer Kalayci, MD1
Abstract: Ang reactive oxygen species (ROS) gihimo sa mga buhing organismo isip resulta sa normal nga metabolismo sa cellular ug mga hinungdan sa palibot, sama sa mga pollutant sa hangin o aso sa sigarilyo. Ang ROS kay reaktibo kaayo nga mga molekula ug makadaot sa mga istruktura sa selula sama sa carbohydrates, nucleic acid, lipid, ug mga protina ug makausab sa ilang mga gimbuhaton. Ang pagbalhin sa balanse tali sa mga oxidant ug mga antioxidant pabor sa mga oxidant gitawag nga �oxidative stress.� Ang regulasyon sa pagkunhod ug pag-oxidizing (redox) nga kahimtang hinungdanon alang sa cell viability, pagpaaktibo, pagdaghan, ug pag-obra sa organ. Ang mga aerobic nga organismo adunay mga integrated antioxidant system, nga naglakip sa enzymatic ug non-enzymatic antioxidants nga kasagaran epektibo sa pagbabag sa makadaot nga mga epekto sa ROS. Bisan pa, sa mga kondisyon sa pathological, ang mga sistema sa antioxidant mahimong mabug-atan. Ang oxidative stress nakatampo sa daghang mga pathological nga kondisyon ug sakit, lakip na ang cancer, neurological disorder, atherosclerosis, hypertension, ischemia/perfusion, diabetes, acute respiratory distress syndrome, idiopathic pulmonary fibrosis, chronic obstructive pulmonary disease, ug asthma. Sa kini nga pagrepaso, among gisumada ang cellular oxidant ug antioxidant nga mga sistema ug hisgutan ang mga epekto sa cellular ug mga mekanismo sa stress sa oxidative.
Ang reactive oxygen species (ROS) gihimo sa mga buhing organismo isip resulta sa normal nga metabolismo sa selula. Sa ubos ngadto sa kasarangang konsentrasyon, naglihok sila sa mga proseso sa pisyolohikal nga selula, apan sa taas nga konsentrasyon, nagpatungha kini ug dili maayong mga kausaban sa mga sangkap sa selula, sama sa lipid, protina, ug DNA.1â6 Ang pagbalhin sa balanse tali sa oxidant/antioxidant pabor sa mga oxidant gitawag nga �oxidative stress.� Ang oxidative stress makatampo sa daghang pathological nga kondisyon, lakip na ang cancer, neurological disorders,7�10 atherosclerosis, hypertension, ischemia/perfusion,11�14 diabetes, acute respiratory distress syndrome, idiopathic pulmonary fibrosis, chronic obstructive pulmonary disease ,15 ug asthma.16–21 Ang mga aerobic nga organismo adunay integrated antioxidant system,� nga naglakip sa enzymatic ug nonenzymatic antioxidants nga kasagaran epektibo sa pagbabag sa makadaot nga mga epekto sa ROS. Bisan pa, sa mga kondisyon sa pathological, ang mga sistema sa antioxidant mahimong mabug-atan. Sa kini nga pagrepaso, among gisumada ang cellular oxidant ug antioxidant nga mga sistema ug regulasyon sa pagkunhod ug pag-oxidizing (redox) nga estado sa kahimsog ug sakit nga estado.
MGA OXIDANTS
Endogenous nga mga Tinubdan sa ROS
Ang ROS gihimo gikan sa molecular oxygen isip resulta sa normal nga metabolismo sa cellular. Ang ROS mahimong bahinon sa 2 ka grupo: free radicals ug nonradicals. Ang mga molekula nga adunay usa o daghan pa nga wala magkapares nga mga electron ug sa ingon naghatag reaktibo sa molekula gitawag nga mga libre nga radikal. Kung ang 2 nga mga libre nga radikal nakigbahin sa ilang mga wala gipares nga mga electron, ang mga nonradical nga porma gihimo. Ang 3 ka dagkong ROS nga adunay pisyolohikal nga kahulogan mao ang superoxide anion (O22.), hydroxyl radical (OH), ug hydrogen peroxide (H2O2). Ang ROS gisumada sa Talaan 1.
Ang superoxide anion naporma pinaagi sa pagdugang sa 1 ka electron sa molekular nga oxygen.22 Kini nga proseso gipataliwala sa nicotine adenine dinucleotide phosphate [NAD(P)H] oxidase o xanthine oxidase o sa mitochondrial electron transport system. Ang nag-unang dapit alang sa paghimo sa superoxide anion mao ang mitochondria, ang makinarya sa selula sa paghimo sa adenosine triphosphate. Kasagaran, ang mga electron gibalhin pinaagi sa mitochondrial electron transport chain alang sa pagkunhod sa oksiheno ngadto sa tubig, apan gibana-bana nga 1 ngadto sa 3% sa tanang mga electron ang mogawas gikan sa sistema ug makahimo og superoxide. Ang NAD(P)H oxidase makita sa polymorphonuclear leukocytes, monocytes, ug macrophage. Sa phagocytosis, kini nga mga selyula nagpatunghag usa ka pagbuto sa superoxide nga motultol sa kalihokan sa bactericidal. Ang superoxide nakabig ngadto sa hydrogen peroxide pinaagi sa aksyon sa superoxide dismutase (SODs, EC 1.15.1.1). Ang hydrogen peroxide dali nga mokaylap sa lamad sa plasma. Ang hydrogen peroxide giprodyus usab sa xanthine oxidase, amino acid oxidase, ug NAD(P)H oxidase23,24 ug sa mga peroxisome pinaagi sa pagkonsumo sa molekular nga oxygen sa metabolic reactions. Sa sunodsunod nga mga reaksyon nga gitawag ug Haber Weiss ug Fenton nga mga reaksyon, ang H2O2 mahimong mabungkag ngadto sa OH2 sa presensya sa transmission metal sama sa Fe21 o Cu21.25.
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 + �H2O2�?Fe3 +�OH�+ .OH Fenton reaksyon
Ang O 2 mismo maka-react usab sa H2 O2 ug makamugna og OH.26,27 Ang hydroxyl radical mao ang pinaka-reaktibo sa ROS ug makadaot sa mga protina, lipid, ug carbohydrates ug DNA. Makasugod usab kini sa lipid peroxidation pinaagi sa pagkuha og electron gikan sa polyunsaturated fatty acids.
Ang mga granulocytic enzymes dugang nga nagpalapad sa reaktibidad sa H2O2 pinaagi sa eosinophil peroxidase ug myeloperoxidase (MPO). Sa activate neutrophils, H2O2 gigamit sa MPO. Sa presensya sa chloride ion, ang H2O2 nakabig ngadto sa hypochlorous acid (HOCl). Ang HOCl kay oxidative kaayo ug adunay importante nga papel sa pagpatay sa mga pathogens sa mga agianan sa hangin.28 Bisan pa, ang HOCl mahimo usab nga mo-react sa DNA ug mag-aghat sa mga interaksyon sa protina sa DNA ug makahimo og pyrimidine oxidation nga mga produkto ug makadugang sa chloride sa mga base sa DNA.29,30 Eosinophil peroxidase ug MPO usab makatampo sa oxidative stress pinaagi sa pagbag-o sa mga protina pinaagi sa halogenations, nitration, ug protina cross-links pinaagi sa tyrosyl radicals.31-33
Ang ubang mga free radical nga nakuha sa oxygen mao ang peroxyl radicals (ROO$). Ang pinakasimple nga porma niini nga mga radical mao ang hydro-peroxyl radical (HOO$) ug adunay papel sa fatty acid peroxidation. Ang mga libreng radicals mahimong mag-trigger sa lipid peroxidation chain reactions pinaagi sa pagkuha sa hydrogen atom gikan sa side-chain methylene carbon. Ang lipid radical dayon mo-react sa oxygen aron makahimo og peroxyl radical. Ang peroxyl radical nagpasiugda sa kadena nga reaksyon ug nag-usab sa polyunsaturated fatty acids ngadto sa lipid hydroperoxides. Ang mga lipid hydroperoxide dili kaayo lig-on ug dali nga madunot sa mga sekondaryang produkto, sama sa aldehydes (sama sa 4-hydroxy-2,3-nonenal) ug malondialdehydes (MDAs). Ang Isoprostanes maoy laing grupo sa mga produkto sa lipid peroxidation nga namugna pinaagi sa peroxidation sa arachidonic acid ug nakit-an usab nga nataas sa plasma ug condensates sa gininhawa sa mga asthmatics.34,35 Ang peroxidation sa mga lipid makasamok sa integridad sa mga membrana sa selula ug mosangpot sa pag-usab sa istruktura sa lamad.
Ang hydrogen peroxide, superoxide radical, oxidized glutathione (GSSG), MDAs, isoprostanes, carbonyls, ug nitrotyrosine daling masukod gikan sa plasma, dugo, o bronchoalveolar lavage samples isip biomarker sa oxidation pinaagi sa standardized assays.
Exogenous nga Tinubdan sa mga Oksiheno
Aso sa Sigarilyo
Ang aso sa sigarilyo adunay daghang mga oxidant ug free radicals ug organic compounds, sama sa superoxide ug nitric oxide.36 Dugang pa, ang inhalation sa aso sa sigarilyo ngadto sa baga nagpalihok usab sa pipila ka endogenous nga mga mekanismo, sama sa pagtipon sa neutrophils ug macrophage, nga dugang nga nagdugang sa kadaot sa oxidant. .
Pagkaladlad sa Ozone
Ang pagkaladlad sa ozone mahimong hinungdan sa peroxidation sa lipid ug mag-aghat sa pag-agos sa mga neutrophil ngadto sa epithelium sa agianan sa hangin. Ang hamubo nga pagkaladlad sa ozone hinungdan usab sa pagpagawas sa mga tigpataliwala sa pagpanghubag, sama sa MPO, eosinophil cationic protein ug usab lactate dehydrogenase ug albumin.37 Bisan sa himsog nga mga hilisgutan, ang pagkaladlad sa ozone hinungdan sa pagkunhod sa mga function sa pulmonary.38 Gipakita ni Cho et al39 nga particulate matter (pagsagol sa solid nga mga partikulo ug mga tinulo nga likido nga gisuspinde sa hangin) nag-catalyze sa pagkunhod sa oxygen.
Hyperoxia
Ang hyperoxia nagtumong sa mga kondisyon nga mas taas ang lebel sa oksiheno kay sa normal nga partial pressure sa oxygen sa baga o ubang mga tisyu sa lawas. Kini modala ngadto sa mas dako nga produksyon sa reactive oxygen ug nitrogen species.40,41
Ionizing Radiation
Ang ionizing radiation, sa presensya sa O2, nag-convert sa hydroxyl radical, superoxide, ug organic radicals ngadto sa hydrogen peroxide ug organic hydroperoxides. Kini nga mga hydroperoxide nga mga espisye mo-react sa redox active metal ions, sama sa Fe ug Cu, pinaagi sa Fenton reactions ug sa ingon nag-aghat sa oxidative stress.42,43 Narayanan et al44 nagpakita nga ang mga fibroblast nga na-expose sa alpha particles adunay dakong pagtaas sa intracellular O2 2. ug H2O2 produksyon pinaagi sa plasma membrane-bound NADPH oxidase.44 Signal transduction molecules, sama sa extracellular signal-regulated kinase 1 ug 2 (ERK1/2), c-Jun N-terminal kinase (JNK), ug p38, ug transcription factors, sama sa activator protein-1 (AP-1), nuclear factor-kB (NF-kB), ug p53, gi-activate, nga moresulta sa pagpahayag sa radiation response’related genes.45-50 Ultraviolet A (UVA) nga mga photon nag-trigger sa oxidative reactions pinaagi sa excitation sa endogenous photosensitizers, sama sa porphyrins, NADPH oxidase, ug riboflavin. Ang 8-Oxo-7,8- dihydroguanine (8-oxoGua) mao ang nag-unang UVA-mediated DNA oxidation nga produkto nga naporma pinaagi sa oksihenasyon sa OH radical, 1-electron oxidants, ug singlet oxygen nga kasagarang mo-react sa guanine.51 Ang pagporma sa guanine radical cation sa nahilit nga DNA gipakita nga episyenteng mahitabo pinaagi sa direktang epekto sa ionizing radiation.52,53 Human sa exposure sa ionizing radiation, ang intracellular level sa glutathione (GSH) mikunhod sulod sa hamubo nga termino apan unya misaka pag-usab.54
Bug-at nga Metal Ion
Ang mabug-at nga metal nga mga ion, sama sa iron, copper, cadmium, mercury, nickel, lead, ug arsenic, mahimong mag-aghat sa pagmugna og mga reactive radicals ug hinungdan sa pagkadaot sa cellular pinaagi sa pagkunhod sa mga kalihokan sa enzyme pinaagi sa lipid peroxidation ug reaksyon sa nuclear proteins ug DNA.55
Usa sa labing importante nga mga mekanismo sa metal-mediated free radical generation mao ang pinaagi sa Fenton-type nga reaksyon. Ang superoxide ion ug hydrogen peroxide mahimong makig-interact sa transisyon nga mga metal, sama sa iron ug copper, pinaagi sa metal catalyzed Haber Weiss/Fenton reaction aron maporma ang OH radicals.
Gawas sa Fenton-type ug HaberWeiss-type nga mga mekanismo, ang pipila ka mga metal nga ion mahimong direktang mo-react sa mga molekula sa cellular aron makamugna og mga free radical, sama sa thiol radical, o mag-aghat sa cell signaling pathways. Kini nga mga radical mahimo usab nga mo-react sa ubang mga thiol molekula aron makamugna og O22.. O22. nakabig ngadto sa H2O2, nga maoy hinungdan sa dugang nga oxygen radical generation. Ang ubang mga metal, sama sa arsenite, nag-aghat sa pagporma sa ROS sa dili direkta pinaagi sa pagpaaktibo sa mga radikal nga sistema sa paghimo sa mga selula.56
Ang arsenic kay makahilo kaayo nga elemento nga naggama ug lain-laing ROS, apil ang superoxide (O2 2), singlet oxygen (1O2), peroxyl radical (ROO ), nitric oxide (NO ), hydrogen peroxide (H2O2), ug dimethylarsinic peroxyl radicals [( CH3)2AsOO ].57�59 Arsenic (III) compounds makapugong sa antioxidant enzymes, ilabina sa GSH-dependent enzymes, sama sa glutathione-S-transferases (GSTs), glutathione peroxidase (GSH-Px), ug GSH reductase, pinaagi sa bind - sa ilang sulfhydryl (�SH) nga mga grupo.60,61
Ang tingga nagdugang sa lipid peroxidation.62 Mahinungdanon nga pagkunhod sa kalihokan sa tissue SOD ug utok GPx ang gitaho pagkahuman sa pagkaladlad sa tingga.63,64 Ang pag-ilis sa zinc, nga nagsilbi nga cofactor sa daghang mga enzyme pinaagi sa tingga, nagdala sa pagka-aktibo sa ingon nga mga enzyme. Ang pagkaladlad sa tingga mahimong hinungdan sa pagdili sa GST pinaagi sa pag-apekto sa mga thiol sa tisyu.
Ang ROS nga namugna sa metal-catalyzed nga mga reaksyon makausab sa mga base sa DNA. Tulo ka base substitutions, G / C, G / T, ug C / T, mahimong mahitabo ingon sa usa ka resulta sa oxidative kadaot sa metal ions, sama sa Fe21, Cu21, ug Ni21. Gipakita ni Reid et al65 nga ang G / C kasagarang gihimo sa Fe21 samtang ang pagpuli sa C / T sa Cu21 ug Ni21.
MGA ANTIOXIDANTS
Ang lawas sa tawo nasangkapan sa lainlaing mga antioxidant nga nagsilbi aron mabalanse ang epekto sa mga oxidant. Alang sa tanan nga praktikal nga katuyoan, kini mahimong bahinon sa 2 nga mga kategorya: enzymatic (Table 2) ug nonenzymatic (Table 3).
Enzymatic Antioxidants
Ang mga mayor nga enzymatic antioxidants sa baga mao ang SODs (EC 1.15.1.11), catalase (EC 1.11.1.6), ug GSH-Px (EC 1.11.1.9). Dugang pa niining mga mayor nga enzyme, ang ubang mga antioxidant, lakip ang heme oxygenase-1 (EC 1.14.99.3), ug redox proteins, sama sa thioredoxins (TRXs, EC 1.8.4.10), peroxiredoxins (PRXs, EC 1.11.1.15), ug glutaredoxins , nakit-an usab nga adunay hinungdanon nga papel sa mga depensa sa pulmonary antioxidant.
Tungod kay ang superoxide mao ang nag-unang ROS nga gihimo gikan sa lain-laing mga tinubdan, ang dismutation niini pinaagi sa SOD mao ang labing importante alang sa matag cell. Ang tanan nga 3 ka porma sa SOD, nga mao, CuZn-SOD, Mn-SOD, ug EC-SOD, kaylap nga gipahayag sa baga sa tawo. Ang Mn-SOD na-localize sa mitochondria matrix. Ang EC-SOD panguna nga na-localize sa extracellular matrix, labi na sa mga lugar nga adunay taas nga kantidad sa type I collagen fibers ug sa palibot sa pulmonary ug systemic nga mga sudlanan. Nakit-an usab kini sa bronchial epithelium, alveolar epithelium, ug alveolar macrophage.66,67 Sa kinatibuk-an, ang CuZn-SOD ug Mn-SOD sa kasagaran gihunahuna nga molihok isip bulk scavengers sa superoxide radicals. Ang medyo taas nga lebel sa EC-SOD sa baga nga adunay espesipikong pagbugkos sa mga sangkap sa extracellular matrix mahimong magrepresentar sa usa ka sukaranan nga sangkap sa proteksyon sa matrix sa baga.68
Ang H2O2 nga gihimo sa aksyon sa mga SOD o ang aksyon sa mga oxidases, sama sa xanthine oxidase, gipaubos sa tubig pinaagi sa catalase ug ang GSH-Px. Ang Catalase anaa isip usa ka tetramer nga gilangkuban sa 4 ka managsama nga monomer, nga ang matag usa adunay usa ka heme nga grupo sa aktibo nga dapit. Ang pagkadaot sa H2O2 nahimo pinaagi sa pagkakabig tali sa 2 ka conformation sa catalase-ferricatalase (iron coordinated to water) ug compound I (iron complexed with oxygen atom). Ang Catalase usab nagbugkos sa NADPH isip usa ka pagkunhod nga katumbas aron mapugngan ang oxidative inactivation sa enzyme (pagporma sa compound II) pinaagi sa H2O2 ingon nga kini gipaubos sa tubig.69
Ang mga enzyme sa redox cycle nga responsable sa pagkunhod sa H2O2 ug lipid hydroperoxides (namugna isip resulta sa membrane lipid peroxidation) naglakip sa GSH-Pxs.70 Ang GSH-Pxs usa ka pamilya sa tetrameric enzymes nga adunay talagsaon nga amino acid selenocysteine sulod sa aktibo nga mga site ug mogamit og mga low-molecular-weight nga thiols, sama sa GSH, aron makunhuran ang H2O2 ug lipid peroxide ngadto sa katugbang nga mga alkohol niini. Upat ka GSH-Pxs ang gihulagway, gi-encode sa lain-laing mga gene: GSH-Px-1 (cellular GSH-Px) kay ubiquitous ug makapamenos sa H2O2 ug fatty acid peroxides, apan dili esterified peroxyl lipids.71 Esterified lipids gipakunhod sa membrane-bound GSH -Px-4 (phospholipid hydroperoxide GSH-Px), nga makagamit ug ubay-ubay nga lainlain nga mga low-molecular-weight nga thiols isip mga katumbas nga pagkunhod. Ang GSH-Px-2 (gastrointestinal GSH-Px) na-localize sa gastrointestinal epithelial cells diin nagsilbi kini nga pagpakunhod sa dietary peroxides.72 GSH-Px-3 (extracellular GSH-Px) mao lamang ang miyembro sa GSH-Px nga pamilya nga nagpuyo sa ang extracellular compartment ug gituohan nga usa sa pinaka importante nga extracellular antioxidant enzyme sa mammals. Niini, ang extracellular GSH-Px ang labing kaylap nga gisusi sa baga sa tawo.73
Dugang pa, ang paglabay sa H2O2 suod nga nalangkit sa ubay-ubay nga thiol-containing enzymes, nga mao, TRXs (TRX1 ug TRX2), thioredoxin reductases (EC 1.8.1.9) (TRRs), PRXs (nga thioredoxin peroxidases), ug glutaredoxins.74
Duha ka TRX ug TRR ang gihulagway sa mga selula sa tawo, nga anaa sa cytosol ug mitochondria. Sa baga, ang TRX ug TRR gipahayag sa bronchial ug alveolar epithelium ug macrophage. Unom ka lainlaing PRX ang nakit-an sa mga selula sa tawo, nga lahi sa ilang ultrastructural compartmentalization. Gipadayag sa mga eksperimento nga pagtuon ang kamahinungdanon sa PRX VI sa pagpanalipod sa alveolar epithelium. Ang baga sa tawo nagpahayag sa tanang PRXs sa bronchial epithelium, alveolar epithelium, ug macrophage.75 Ang PRX V bag-o lang nakit-an nga naglihok isip usa ka peroxynitrite reductase, 76 nga nagpasabot nga kini mahimong molihok isip usa ka potensyal nga protective compound sa pagpalambo sa ROS-mediated lung injury .77
Kasagaran sa kini nga mga antioxidant mao ang kinahanglanon sa NADPH ingon usa ka pagkunhod nga katumbas. Ang NADPH nagmintinar sa catalase sa aktibong porma ug gigamit isip cofactor sa TRX ug GSH reductase (EC 1.6.4.2), nga nag-convert sa GSSG ngadto sa GSH, usa ka co-substrate alang sa GSH-Pxs. Ang intracellular NADPH, sa baylo, namugna pinaagi sa pagkunhod sa NADP1 pinaagi sa glucose-6-phosphate dehydrogenase, ang una ug rate-limiting enzyme sa pentose phosphate nga agianan, sa panahon sa pagkakabig sa glucose-6-phosphate ngadto sa 6-phosphogluconolactone. Pinaagi sa pagmugna sa NADPH, ang glucose-6-phosphate dehydrogenase usa ka kritikal nga determinant sa cytosolic GSH buffering capacity (GSH/GSSG) ug, busa, mahimong isipon nga usa ka importante, regulatory antioxidant enzyme.78,79
GSTs (EC 2.5.1.18), laing antioxidant enzyme nga pamilya, dili aktibo sa secondary metabolites, sama sa unsaturated aldehydes, epoxides, ug hydroperoxides. Tulo ka dagkong pamilya sa GST ang gihulagway: cytosolic GST, mitochondrial GST,80,81 ug membrane-associated microsomal GST nga adunay papel sa eicosanoid ug GSH metabolism.82 Pito ka klase sa cytosolic GST ang giila sa mammalian, gitudlo nga Alpha, Mu, Pi, Sigma, Theta, Omega, ug Zeta.83-86 Atol sa dili-stressed nga mga kondisyon, ang klase Mu ug Pi GST nakig-interact sa kinases Ask1 ug JNK, matag usa, ug nagpugong niini nga mga kinase.87'89 Gipakita nga ang GSTP1 nagbulag gikan sa Ang JNK agig tubag sa oxidative stress.89 Ang GSTP1 pisikal usab nga nakig-uban sa PRX VI ug nagdala ngadto sa pagkaayo sa PRX enzyme nga kalihokan pinaagi sa glutathionylation sa oxidized protein.90
Nonenzymatic Antioxidants
Ang nonenzymatic antioxidants naglakip sa ubos nga molekular-timbang nga mga compound, sama sa bitamina (bitamina C ug E), b-carotene, uric acid, ug GSH, usa ka tripeptide (Lg-glutamyl-L-cysteinyl-L-glycine) nga naglangkob sa usa ka thiol ( sulfhydryl) nga grupo.
Bitamina C (Ascorbic Acid)
Ang matunaw sa tubig nga bitamina C (ascorbic acid) naghatag sa intracellular ug extracellular aqueous-phase nga kapasidad nga antioxidant sa panguna pinaagi sa pag-scavenging sa oxygen free radicals. Gibag-o niini ang mga free radical sa bitamina E balik sa bitamina E. Ang lebel sa plasma niini gipakita nga nagkunhod sa edad.91,92
Bitamina E (a-Tocopherol)
Ang matunaw nga lipid nga bitamina E gikonsentrar sa hydrophobic interior site sa cell membrane ug mao ang panguna nga depensa batok sa kadaot sa lamad nga gipahinabo sa oxidant. Ang Vitamin E nagdonar og electron sa peroxyl radical, nga gihimo atol sa lipid peroxidation. Ang a-Tocopherol mao ang labing aktibo nga porma sa bitamina E ug ang mayor nga lamad nga gigapos nga antioxidant sa selula. Ang Vitamin E nag-trigger sa apoptosis sa mga selula sa kanser ug nagpugong sa mga free radical formations.93
Glutathione
Ang GSH kay daghan kaayo sa tanang cell compartment ug mao ang mayor nga matunaw nga antioxidant. GSH/GSSG ratio mao ang usa ka mayor nga determinant sa oxidative stress. Gipakita sa GSH ang iyang antioxidant nga mga epekto sa daghang paagi.94 Kini nag-detoxify sa hydrogen peroxide ug lipid peroxide pinaagi sa aksyon sa GSH-Px. Gidonar sa GSH ang iyang electron sa H2O2 aron makunhuran kini ngadto sa H2O ug O2. Ang GSSG gipaubos na usab ngadto sa GSH pinaagi sa GSH reductase nga naggamit sa NAD(P)H isip electron donor. Importante usab ang GSH-Pxs alang sa pagpanalipod sa cell membrane gikan sa lipid peroxidation. Ang pagkunhod sa glutathione nagdonar sa mga proton sa mga lipid sa lamad ug nanalipod kanila gikan sa mga pag-atake sa oxidant.95
Ang GSH usa ka cofactor alang sa daghang mga detoxifying enzyme, sama sa GSH-Px ug transferase. Kini adunay papel sa pagbag-o sa bitamina C ug E balik sa ilang aktibo nga porma. Gipanalipdan sa GSH ang mga selula batok sa apoptosis pinaagi sa pagpakig-uban sa proapoptotic ug antiapoptotic signaling pathways.94 Kini usab nag-regulate ug nagpalihok sa daghang mga transcription nga mga hinungdan, sama sa AP-1, NF-kB, ug Sp-1.
Carotenoids (b-Carotene)
Ang mga carotenoid mga pigment nga makita sa mga tanum. Sa panguna, ang b-carotene nakit-an nga mo-react sa peroxyl (ROO), hydroxyl (OH), ug superoxide (O22.) radicals.96 Ang mga carotenoid nagpakita sa ilang antioxidant nga mga epekto sa ubos nga oxygen partial pressure apan mahimong adunay pro-oxidant nga epekto sa mas taas nga oxygen. konsentrasyon.97 Ang duha ka carotenoids ug retinoic acids (RAs) makahimo sa pag-regulate sa transcription factors.98 b-Carotene mopugong sa oxidant-induced NF-kB activation ug interleukin (IL)-6 ug tumor necrosis factor-a production. Ang mga carotenoid usab makaapekto sa apoptosis sa mga selula. Ang mga antiproliferative nga epekto sa RA gipakita sa daghang mga pagtuon. Kini nga epekto sa RA gipataliwala sa panguna sa mga receptor sa retinoic acid ug lainlain sa mga tipo sa selula. Sa mammary carcinoma cells, ang retinoic acid receptor gipakita nga maka-trigger sa growth inhibition pinaagi sa pag-aghat sa cell cycle arrest, apoptosis, o pareho.99,100
ANG EPEKTO SA OXIDATIVE STRESS: GENETIC, PHYSIOLOGICAL, & BIOCHEMICAL MECHANISMS
Ang oxidative stress mahitabo kung ang balanse tali sa antioxidants ug ROS mabalda tungod sa pagkahurot sa mga antioxidant o pagtipon sa ROS. Kung mahitabo ang stress sa oxidative, ang mga selyula mosulay sa pagsumpo sa mga epekto sa oxidant ug ibalik ang balanse sa redox pinaagi sa pagpaaktibo o pagpahilom sa mga gene nga nag-encode sa mga defensive enzyme, transkripsyon nga mga hinungdan, ug istruktura nga protina. sa importante nga determinants sa oxidative stress sa lawas. Ang mas taas nga produksiyon sa ROS sa lawas mahimong magbag-o sa istruktura sa DNA, moresulta sa pagbag-o sa mga protina ug lipid, pagpaaktibo sa daghang mga hinungdan sa transkripsyon nga gipahinabo sa stress, ug paghimo sa mga pro-inflammatory ug anti-inflammatory cytokine.
Mga Epekto sa Oxidative Stress Sa DNA
Ang ROS mahimong mosangpot sa pagbag-o sa DNA sa daghang paagi, nga naglakip sa pagkadaot sa mga base, single-o double-stranded nga DNA break, purine, pyrimidine o sugar-bound modifications, mutation, deletion o translocations, ug cross-linking sa mga protina. Kadaghanan niini nga mga pagbag-o sa DNA (Fig. 1) may kalabutan kaayo sa carcinogenesis, pagtigulang, ug neurodegenerative, cardiovascular, ug autoimmune nga mga sakit. Ang aso sa tabako, redox nga mga metal, ug mga nonredox nga metal, sama sa iron, cadmium, chrome, ug arsenic, nalangkit usab sa carcinogenesis ug pagkatigulang pinaagi sa pagmugna og mga free radical o pagbugkos sa thiol nga mga grupo. Ang pagporma sa 8-OH-G mao ang labing nailhan nga kadaot sa DNA nga nahitabo pinaagi sa oxidative stress ug usa ka potensyal nga biomarker alang sa carcinogenesis.
Ang mga rehiyon sa promoter sa mga gene adunay consensus sequence alang sa transcription factor. Kining mga transcription factor�binding sites adunay GC-rich sequence nga daling maatake sa oxidant. Ang pagporma sa 8-OH-G DNA sa transcription factor binding sites makausab sa pagbugkos sa transcription factors ug sa ingon mausab ang ekspresyon sa mga may kalabutan nga mga gene sama sa gipakita sa AP-1 ug Sp-1 nga target sequences.103 Gawas sa 8-OH-G, Ang 8,59 -cyclo-29 -deoxyadenosine (cyclo-dA) gipakita usab nga makapugong sa transkripsyon gikan sa usa ka reporter gene sa usa ka sistema sa selula kon anaa sa kahon sa TATA.104 Ang TATA-binding protein nagpasiugda sa transkripsyon pinaagi sa pag-usab sa bending sa DNA . Ang pagbugkos sa TATA-binding protein mahimong madaot tungod sa presensya sa cyclo-dA.
Ang oxidative stress hinungdan sa pagkawalay kalig-on sa microsatellite (mubo nga tandem repeats) nga mga rehiyon. Ang redox active metal ions, hydroxyl radicals nagdugang sa microsatellite instability.105 Bisan tuod ang single-stranded DNA break tungod sa oxidant injury daling maagwanta sa mga selula, ang double-stranded DNA break nga gipahinabo sa ionizing radiation mahimong dakong hulga sa cell survival.106
Ang methylation sa mga isla sa CpG sa DNA usa ka importante nga mekanismo sa epigenetic nga mahimong moresulta sa pagpahilom sa gene. Ang oxidation sa 5-MeCyt ngadto sa 5-hydroxymethyl uracil (5-OHMeUra) mahimong mahitabo pinaagi sa deamination/oxidation reactions sa thymine o 5-hydroxymethyl cytosine intermediates.107 Dugang pa sa modulating gene expression, ang DNA methylation daw makaapekto usab sa chromatin organization.108 Ang aberrant DNA methylation patterns tungod sa oxidative attacks makaapekto usab sa DNA repair activity.
Mga Epekto Sa Oxidative Stress Sa Lipid
Ang ROS makaaghat sa lipid peroxidation ug makabalda sa lamad nga lipid bilayer nga kahikayan nga mahimong dili aktibo sa membrane-bound receptors ug enzymes ug makadugang sa tissue permeability.109 Ang mga produkto sa lipid peroxidation, sama sa MDA ug unsaturated aldehydes, makahimo sa pagpa-inactivate sa daghang cellular proteins pinaagi sa pagporma sa protina cross -linkages.110�112 4-Hydroxy-2-nonenal hinungdan sa pagkahurot sa intracellular GSH ug pag-aghat sa produksiyon sa peroxide,113,114 pagpaaktibo sa epidermal growth factor receptor,115 ug pagduso sa produksiyon sa fibronectin.116 Lipid peroxidation nga mga produkto, sama sa isoprostanes ug thiobarbituric acid reactive substances , gigamit isip dili direkta nga biomarker sa oxidative stress, ug ang pagtaas sa lebel gipakita sa exhaled breath condensate o bronchoalveolar lavage fluid o baga sa chronic obstructive pulmonary disease nga mga pasyente o mga hinabako.117-119
Mga Epekto sa Oxidative Stress sa Protina
Ang ROS mahimong hinungdan sa pagkabahin sa kadena sa peptide, pagbag-o sa elektrikal nga karga sa mga protina, cross-linking sa mga protina, ug oksihenasyon sa piho nga mga amino acid ug busa modala ngadto sa dugang nga pagkadaling maapektuhan sa proteolysis pinaagi sa pagkadaut sa piho nga mga protease.120 Ang mga residu sa cysteine ug methionine sa mga protina ilabina nga mas daling madala sa oksihenasyon.121 Ang oxidation sa sulfhydryl groups o methionine residues sa mga protina maoy hinungdan sa conformational change, protein unfolding, ug degradation.8,121�123 Ang mga enzyme nga adunay mga metal sa o duol sa ilang aktibo nga mga dapit ilabinang mas sensitibo sa metal catalyzed oxidation. Ang oxidative nga pagbag-o sa mga enzyme gipakita nga makapugong sa ilang mga kalihokan.124,125
Sa pipila ka mga kaso, ang piho nga oksihenasyon sa mga protina mahimong mahitabo. Pananglitan, ang methionine mahimong oxidized methionine sulfoxide126 ug phenylalanine ngadto sa o-tyrosine127; Ang mga grupo sa sulfhydryl mahimong ma-oxidized aron maporma ang mga disulfide bond; 128 ug ang mga grupo sa carbonyl mahimong ipasok sa kilid nga kadena sa mga protina. Ang gamma rays, metal-catalyzed oxidation, HOCl, ug ozone mahimong hinungdan sa pagporma sa carbonyl groups.129
Mga Epekto sa Oxidative Stress sa Signal Transduction
Ang ROS makahimo sa pagpahayag sa daghang mga gene nga nalambigit sa signal transduction.1,130 Ang taas nga ratio sa GSH/GSSG importante alang sa pagpanalipod sa selula gikan sa oxidative nga kadaot. Ang pagkabalda niini nga ratio maoy hinungdan sa pagpaaktibo sa redox sensitive transcription nga mga hinungdan, sama sa NF-kB, AP-1, nukleyar nga hinungdan sa mga activate nga T cells ug hypoxia-inducible factor 1, nga nalangkit sa makapahubag nga tubag. Ang pagpaaktibo sa mga hinungdan sa transkripsyon pinaagi sa ROS makab-ot pinaagi sa mga signal transduction cascades nga nagpadala sa kasayuran gikan sa gawas hangtod sa sulod sa cell. Tyrosine kinase receptors, kadaghanan sa growth factor receptors, sama sa epidermal growth factor receptor, vascular endothelial growth factor receptor, ug receptor alang sa platelet-derived growth factor, protein tyrosine phosphatases, ug serine/threonine kinases maoy mga target sa ROS.131-133 Ang extra-cellular signal-regulated kinases, JNK, ug p38, nga mga membro sa mitogen-activated protein kinase family ug nalambigit sa daghang proseso sa cell lakip na ang proliferation, differentiation, ug apoptosis, mahimo usab nga ma-regulate sa mga oxidant.
Ubos sa mga kondisyon sa stress sa oxidative, ang mga residue sa cysteine sa DNA-binding site sa c-Jun, pipila ka mga subunit sa AP-1, ug ang nagpugong nga kB kinase moagi sa mabalik nga S-glutathiolation. Ang Glutaredoxin ug TRX gikataho nga adunay importante nga papel sa regulasyon sa redox-sensitive signaling pathways, sama sa NF-kB ug AP-1, p38 mitogen-activated protein kinase, ug JNK.134�137
Ang NF-kB mahimong ma-activate agig tubag sa oxidative stress nga mga kondisyon, sama sa ROS, free radicals, ug UV irradiation.138 Phosphorylation of IkB frees NF-kB ug gitugotan kini nga mosulod sa nucleus aron ma-activate ang transcription sa gene.139 Daghang kinase ang adunay gitaho sa phosphorylate IkBs sa serine residues. Kini nga mga kinases mao ang mga target sa mga oxidative signal alang sa pagpaaktibo sa NF-kB.140 Ang pagkunhod sa mga ahente nagpalambo sa NF-kB DNA binding, samtang ang oxidizing agents nagpugong sa DNA binding sa NF-kB. Ang TRX mahimong magbuhat ug 2 ka kaatbang nga mga aksyon sa regulasyon sa NF-kB: sa cytoplasm, gibabagan niini ang pagkadaot sa IkB ug gipugngan ang pagpaaktibo sa NF-kB apan gipauswag ang pagbugkos sa DNA sa NF-kB sa nucleus.141 Pagpaaktibo sa NF-kB pinaagi sa pagkadaot nga may kalabotan sa oksihenasyon sa IkB moresulta sa pagpaaktibo sa daghang mga antioxidant nga depensa nga may kalabutan nga mga gene. Ang NF-kB nag-regulate sa pagpahayag sa daghang mga gene nga miapil sa immune response, sama sa IL-1b, IL-6, tumor necrosis factor-a, IL-8, ug daghang mga molekula sa adhesion.142,143 Ang NF-kB usab nag-regulate sa angiogenesis ug pagdaghan ug pagkalahi sa mga selula.
Ang AP-1 gi-regulate usab sa redox state. Sa presensya sa H2O2, ang pipila ka mga ion nga metal mahimong mag-aghat sa pagpaaktibo sa AP-1. Ang pagtaas sa ratio sa GSH / GSSG nagpalambo sa AP-1 nga pagbugkos samtang ang GSSG nagpugong sa pagbugkos sa DNA sa AP-1.144 nga pagbugkos sa DNA sa Fos / Jun heterodimer gipataas pinaagi sa pagkunhod sa usa ka gitipigan nga cysteine sa DNA-binding domain sa matag usa ang mga protina, 145 samtang ang DNA binding sa AP-1 mahimong pugngan sa GSSG sa daghang matang sa selula, nga nagsugyot nga ang disulphide bond formation pinaagi sa cysteine residues makapugong sa AP-1 DNA binding.146,147 Signal transduction pinaagi sa oxidative stress gisumada sa Figure 2.
MGA KASABUTAN
Ang tensiyon sa oxidative mahimong mutungha gikan sa sobrang produksiyon sa ROS pinaagi sa mga metabolikong reaksyon nga naggamit ug oksiheno ug nagbalhin sa balanse tali sa oxidant/antioxidant mga kahimtang nga pabor sa mga oxidant. Ang ROS gihimo sa mga kalihokan sa cellular metabolic ug mga hinungdan sa kalikopan, sama sa mga pollutant sa hangin o aso sa sigarilyo. Ang ROS kay reaktibo kaayo nga mga molekula tungod sa wala magkapares nga mga electron sa ilang estraktura ug nag-react sa daghang biological nga macromolecules sa cell, sama sa carbohydrates, nucleic acids, lipids, ug proteins, ug giusab ang ilang mga gimbuhaton. Ang ROS nakaapekto usab sa pagpahayag sa daghang mga gene pinaagi sa pag-upregulation sa redox-sensitive transcription nga mga hinungdan ug chromatin remodeling pinaagi sa pag-usab sa histone acetylation/deacetylation. Ang regulasyon sa redox state hinungdanon alang sa cell viability, activation, proliferation, ug organ function.
mga pakisayran
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals ug antioxidants sa oxidative stress-induced cancer. Chem Biol Interact. 2006;160:1–40.
2. Halliwell B, Gutteridge JMC. Libre nga Radikal sa Biology ug Medisina. ika-3 nga ed. New York: Oxford University Press; 1999.
3. Marnett LJ. Lipid peroxidationdDNA kadaot sa malondialdehyde. Mutat Res. 1999;424:83–95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal formation atol sa ischemia ug reperfusion sa gamay nga tinai sa ilaga. �Kinabuhi Sci. 1995;57:785–789.
5. Stadtman ER. Ang papel sa mga oxidant species sa pagkatigulang. Curr Med Chem. 2004;11:1105–1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Ang lipid peroxidation-induced putative malondialdehyde'DNA adducts sa mga tisyu sa dughan sa tawo. Cancer Epidemiol Biomarkers Prev. 1996;5:705–710.
7. Jenner P. Oxidative stress sa Parkinson's disease. Ann Neurol. 2003;53: S26�S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Usa ka pagsusi sa oxidative nga kadaot sa mga protina, lipid, ug DNA sa utok gikan sa mga pasyente nga adunay Alzheimer's disease. J Neurochem. 1997;68:2061–2069.
9. Sayre LM, Smith MA, Perry G. Chemistry ug biochemistry sa oxidative stress sa neurodegenerative nga sakit. Curr Med Chem. 2001;8:721–738.
10. Toshniwal PK, Zarling EJ. Ebidensya alang sa dugang nga lipid peroxidation sa multiple sclerosis. Ang Neurochem Res. 1992;17:205–207.
11. Dhalla NS, Temsah RM, Netticadan T. Papel sa oxidative stress sa cardiovascular nga mga sakit. J Hypertens. 2000;18:655–673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, et al. Pagtuon sa oxidative stress sa usa ka modelo sa ilaga sa chronic brain hypoperfusion. Neurochem Int. 2005;46:601–611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Ang produksiyon sa superoxide anion nadugangan sa usa ka modelo sa genetic hypertension: papel sa endothelium. Hypertension. 1999;33:1353–1358.
14. Kukreja RC, Hess ML. Ang oxygen free-radical nga sistema: gikan sa mga equation pinaagi sa mga interaksyon sa lamad sa protina hangtod sa kadaot sa cardiovascular ug proteksyon. Cardiovasc Res. 1992;26:641–655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, et al. Ang pagpanigarilyo nagpahinabog pagtaas sa kadaot sa oxidative DNA, 8-hydroxydeoxyguanosine, sa sentro nga lugar sa baga sa tawo. Carcinogenesis. 1997;18:1763–1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Oxidative ug nitrosative nga mga panghitabo sa hika. Libre nga Radic Biol Med. 2003;35:213–225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. Correlation sa systemic superoxide dismutase deficiency sa airflow obstruction sa hika. Am J Respir Crit Care Med. 2005;172:306–313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, et al. Superoxide dismutase inactivation sa pathophysiology sa asthmatic airway remodeling ug reactivity. Si J Pathol. 2005;166:663–674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Oxidative stress ug ang mga determinant niini sa mga agianan sa hangin sa mga bata nga adunay hika. Alerdyi. 2008;63:1605–1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, et al. Oxidative stress ug genetic ug epidemiologic determinants sa oxidant injury sa childhood asthma. J Allergy Clin Immunol. 2006;118:1097–1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Grabi nga Asthma Research Program. Ang airway glutathione homeostasis giusab sa mga bata nga adunay grabe nga hika: ebidensya sa stress sa oxidant. J Allergy Clin Immunol. 2009;123:146–152.
22. Miller DM, Buettner GR, Aust SD. Transition metals isip catalysts sa "autoxidation" nga mga reaksyon. Libre nga Radic Biol Med. 1990;8:95–108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D�me D, Pommier J. Mekanismo sa hydrogen peroxide formation nga gi-catalyzed sa NADPH oxidase sa thyroid plasma membrane. J Biol Chem. 1991;266:3739–3743.
24. Granger DN. Papel sa xanthine oxidase ug granulocytes sa kadaot sa ischemiareperfusion. Am J Physiol. 1988;255:H1269�H1275.
25. Fenton HJH. Oxidation sa tartaric acid sa atubangan sa puthaw. Si J Chem Soc. 1984;65:899–910.
26. Haber F, Weiss JJ. Ang catalytic decomposition sa hydrogen peroxide pinaagi sa iron salts. Proc R Soc Lond Ser A. 1934;147:332�351.
27. Liochev SI, Fridovich I. Ang Haber Weiss nagbisikleta 70 ka tuig ang milabay: usa ka alternatibong panglantaw. Redox Rep. 2002;7:55—57.
28. Klebanoff SJ. Myeloperoxidase: higala ug kaaway. J Leukoc Biol. 2005;77:598–625.
29. Whiteman M, Jenner A, Halliwell B. Hypochlorous acid-induced base modifications sa nahilit nga calf thymus DNA. Chem Res Toxicol. 1997;10:1240–1246.
30. Kulcharyk PA, Heinecke JW. Ang hypochlorous acid nga gihimo sa myeloperoxidase system sa mga phagocytes sa tawo nag-aghat sa mga covalent cross-link tali sa DNA ug protina. Biochemistry. 2001;40:3648–3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W, et al. Usa ka sugilanon sa duha ka kontrobersiya: ang paghubit sa papel sa peroxidases sa nitrotyrosine formation sa vivo gamit ang eosinophil peroxidase ug myeloperoxidasedeficient nga mga ilaga, ug ang kinaiya sa peroxidase-generated reactive nitrogen species. J Biol Chem. 2002;277:17415–17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, et al. Ang halapad nga eosinophil degranulation ug peroxidase-mediated oxidation sa airway proteins dili mahitabo sa usa ka mouse ovalbumin-challenge model sa pulmonary inflammation. J Immunol. 2001;167:1672–1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Nitrite isip substrate ug tigpugong sa myeloperoxidase. Mga implikasyon alang sa nitrasyon ug paghimo sa hypochlorous acid sa mga lugar sa panghubag. J Biol Chem. 2000;275:11638–11644.
34. Wood LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Ang lipid peroxidation nga gitino sa plasma isoprostanes adunay kalabotan sa kagrabe sa sakit sa malumo nga hika. Mga lipid. 2000;35:967–974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Dugang nga 8-isoprostane, usa ka timaan sa oxidative stress, sa exhaled condensate sa mga pasyente sa asthma. Am J Respir Crit Care Med. 1999;160:216–220.
36. Simbahan DF, Pryor WA. Free-radical chemistry sa aso sa sigarilyo ug ang makahilo nga implikasyon niini. Panglantaw sa Panglawas sa Kalibutan. 1985;64:111–126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, ug uban pa. Ozone-induced nga panghubag nga gisusi sa sputum ug bronchial lavage fluid gikan sa asthmatics: usa ka bag-ong noninvasive nga himan sa epidemiologic nga mga pagtuon sa polusyon sa hangin ug hika. Libre nga Radic Biol Med. 1999;27:1448–1454.
38. Nightingale JA, Rogers DF, Barnes PJ. Epekto sa inhaled ozone sa exhaled nitric oxide, pulmonary function, ug induced sputum sa normal ug asthmatic nga mga subject. Thorax. 1999;54:1061–1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, et al. Redox nga kalihokan sa airborne particulate matter sa lain-laing mga dapit sa Los Angeles Basin. Kalibutan Res. 2005;99:40–47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Differential induction sa extracellular glutathione peroxidase ug nitric oxide synthase 2 sa mga agianan sa hangin sa himsog nga mga indibidwal nga naladlad sa 100% O (2) o aso sa sigarilyo. Am J Respir Cell Mol Biol. 2000;23:350–354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Oxidant-mediated nga kadaot sa baga sa acute respiratory distress syndrome. Crit Care Med. 1999;27:2028–2030.
42. Biaglow JE, Mitchell JB, Held K. Ang kamahinungdanon sa peroxide ug superoxide sa X-ray nga tubag. Int J Radiat Oncol Biol Phys. 1992;22:665–669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Ang copper ion-mediated sensitization sa nuclear matrix attachment sites sa ionizing radiation. Biochemistry. 1993;32:6214–6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Ang mga partikulo sa alpha nagsugod sa biolohikal nga paggama sa superoxide anion ug hydrogen peroxide sa mga selula sa tawo. Cancer Res. 1997;57:3963–3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Pagkasensitibo sa mga kemikal nga oxidant ug radiation sa CHO cell lines kulang sa oxidative pentose cycle nga kalihokan. Int J Radiat Oncol Biol Phys. 1992;22: 671–675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, et al. Manganese
superoxide dismutase-mediated gene expression sa radiation nga gipahinabo
mapahiangay nga mga tubag. Mol Cell Biol. 2003;23:2362–2378.
47. Azzam EI, de Toledo SM, Spitz DR, Gamay nga JB. Oxidative metabolismo
modulates signal transduction ug micronucleus formation sa bystander
mga selula gikan sa a-particle irradiated normal nga fibroblast sa tawo. Cancer Res.
2002;62:5436–5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Ionizing radiation-induced, mitochondria-dependent nga henerasyon sa reaktibo
oxygen/nitrogen. Cancer Res. 2001;61:3894–3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK nga mga agianan sa
mga tubag sa radiation. Oncogene. 2003;22:5885–5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, et al. Thioredoxin
Ang nukleyar nga translocation ug pakig-uban sa redox factor-1 nagpalihok sa AP-1 transcription factor isip tubag sa ionizing radiation. Cancer Res. 2000;60:6688–6695.
51. Cadet J, Douki T, Gasparutto D, Ravanat JL. Oxidative kadaot sa DNA: pagporma, pagsukod ug biochemical bahin. Mutat Res. 2003;531:5–23.
52. Yokoya A, Cunniffe SM, O'Neill P. Epekto sa hydration sa induction sa strand breaks ug base lesions sa plasmid DNA films pinaagi sa gammaradiation. J Am Chem Soc. 2002;124:8859–8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Mga tubag sa cell ug tissue sa kadaot sa oxidative. Lab Invest. 1993;69:261–274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, et al. Nuclear factor kappa B dependent induction sa gamma glutamylcysteine synthetase pinaagi sa ionizing radiation sa T98G human glioblastoma cells. Libre nga Radic Biol Med. 1998;24:1256–1268.
55. Stohs SJ, Bagchi D. Oxidative mekanismo sa toxicity sa metal ions. Libre nga Radic Biol Med. 1995;18:321–336.
56. Leonard SS, Harris GK, Shi X. Metal-induced oxidative stress ug signal transduction. Libre nga Radic Biol Med. 2004;37:1921–1942.
57. Shi H, Shi X, Liu KJ. Oxidative nga mekanismo sa arsenic toxicity ug carcinogenesis. Mol Cell Biochem. 2004;255:67–78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Usa ka potensyal nga mekanismo alang sa pagkadaot sa nitric oxide formation tungod sa dugay nga oral exposure sa arsenate sa mga rabbits. Free Radic Biol Med.2003;35:102—113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. Ang mga break sa DNAstrand nga gipahinabo sa dimethylarsinic acid, usa ka metabolite sa inorganic nga arsenics, kusog nga gipauswag sa mga radikal nga superoxide anion. Biol Pharm Bull. 1995;18:45–58.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Mga mekanismo nga nagpahipi sa arsenic carcinogenesis: sobrang pagkasensitibo sa mga ilaga nga naladlad sa dili organikong arsenic sa panahon sa pagmabdos. Toxicology. 2004;198:31–38.
61. Schiller CM, Fowler BA, Woods JS. Epekto sa arsenic sa pyruvate dehydrogenase activation. Panglantaw sa Panglawas sa Kalibutan. 1977;19:205–207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Ang pag-apil sa mga libre nga radical sa neurological porphyrias ug pagkahilo sa tingga. Mol Cell Biochem. 1991;103:73–83.
63. Tripathi RM, Raghunath R, Mahapatra S. Ang tingga sa dugo ug ang epekto niini sa lebel sa Cd, Cu, Zn, Fe ug hemoglobin sa mga bata. Kinatibuk-ang Kalibutan sa Sci. 2001;277:161–168.
64. Nehru B, Dua R. Ang epekto sa dietary selenium sa lead neurotoxicity. J Environ Pathol Toxicol Oncol. 1997;16:47–50.
65. Reid TM, Feig DI, Loeb LA. Mutagenesis pinaagi sa metal-induced oxygen radicals. Panglantaw sa Panglawas sa Kalibutan. 1994;102(suppl 3):57–61.
66. Kinnula VL, Crapo JD. Ang superoxide dismutase sa baga ug mga sakit sa baga sa tawo. Am J Respir Crit Care Med. 2003;167:1600–1619.
67. Kinnula VL. Pagprodyus ug pagkadaot sa mga metabolite sa oksiheno sa panahon sa makapahubag nga mga estado sa baga sa tawo. Ang Curr Drug Target nga Makapahubag sa Allergy. 2005;4:465–470.
68. Zelko IN, Mariani TJ, Folz RJ. Superoxide dismutase multigene nga pamilya: usa ka pagtandi sa CuZn-SOD (SOD1), Mn-SOD (SOD2), ug EC-SOD (SOD3) nga mga istruktura sa gene, ebolusyon, ug ekspresyon. Libre nga Radic Biol Med. 2002;33:337–349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Mga mekanismo sa pagpanalipod sa catalase pinaagi sa NADPH. Kinetics ug stoichiometry. J Biol Chem. 1999;274:13908–13914.
70. Floh� L. Glutathione peroxidase. Batakang Kinabuhi Sci. 1988;49:663–668.
71. Arthur JR. Ang glutathione peroxidases. Cell Mol Kinabuhi Sci. 2000;57:1825–1835.
72. Chu FF, Doroshow JH, Esworthy RS. Pagpahayag, kinaiya, ug pag-apod-apod sa tisyu sa usa ka bag-ong cellular selenium-dependent glutathione peroxidase, GSHPx-GI. J Biol Chem. 1993;268:2571–2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Extracellular glutathione peroxidase induction sa asthmatic lungs: ebidensya alang sa redox regulation sa ekspresyon sa human airway epithelial cells. FASEB J. 2001;15:70�78.
74. Gromer S, Urig S, Becker K. Ang sistema sa thioredoxin gikan sa siyensya ngadto sa klinika. Med Res Pin. 2004;24:40–89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P��kk� P, et al. Ang piho nga ekspresyon sa selula sa peroxiredoxins sa baga sa tawo ug sarcoidosis sa pulmonary. Thorax. 2002;57:157–164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, et al. Ang peroxiredoxin 5 sa tawo usa ka peroxynitrite reductase. FEBS Lett. 2004;571:161–165.
77. Holmgren A. Antioxidant function sa thioredoxin ug glutaredoxin system. Antioxid Redox Signal. 2000;2:811–820.
78. Dickinson DA, Forman HJ. Glutathione sa depensa ug pagsenyas: mga leksyon gikan sa gamay nga thiol. Ann NY Acad Sci. 2002;973:488–504.
79. Sies H. Glutathione ug ang papel niini sa cellular functions. Libre nga Radic Biol Med. 1999;27:916–921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Parallel evolutionary pathways alang sa glutathione transferases: istruktura ug mekanismo sa mitochondrial class kappa enzyme rGSTK1-1. Biochemistry. 2004;43:52–61.
81. Robinson A, Huttley GA, Booth HS, Board PG. Ang pagmodelo ug bioinformatics nga mga pagtuon sa human kappa class glutathione transferase nagtagna sa usa ka nobela nga ikatulo nga transferase nga pamilya nga adunay homology sa prokaryotic 2-hydroxychromene-2-carboxylate isomerases. Biochem J. 2004;379:541–552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Komon nga mga bahin sa estruktura sa MAPEGda kaylap nga superfamily sa lamad nga may kalabutan nga mga protina nga adunay hilabihan nga nagkalainlain nga mga gimbuhaton sa eicosanoid ug glutathione metabolism. Protina Sci. 1999;8:689–692.
83. Hayes JD, Pulford DJ. Ang glutathione S-transferase supergene nga pamilya: regulasyon sa GST ug ang kontribusyon sa isoenzymes sa cancer chemoprotection ug drug resistance. Crit Rev Biochem Mol Biol. 1995;30:445–600.
84. Armstrong RN. Istruktura, mekanismo sa catalytic, ug ebolusyon sa glutathione transferases. Chem Res Toxicol. 1997;10:2–18.
85. Hayes JD, McLellan LI. Ang glutathione ug glutathione-dependent nga mga enzyme nagrepresentar sa usa ka co-ordinately regulated nga depensa batok sa oxidative stress. Libre nga Radic Res. 1999;31:273–300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Structure, function ug evolution sa glutathione transferases: mga implikasyon alang sa klasipikasyon sa mga nonmammalian nga mga miyembro sa usa ka karaang enzyme superfamily. Biochem J. 2001;360:1–16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, ug uban pa. Ang Glutathione S-transferase Mu modulates sa stress activated signals pinaagi sa pagsumpo sa apoptosis signal-regulating kinase 1. J Biol Chem. 2001;276:12749–12755.
88. Dorion S, Lambert H, Landry J. Ang pagpaaktibo sa p38 signaling pathway pinaagi sa heat shock naglakip sa dissociation sa glutathione S-transferase Mu gikan sa Ask1. J Biol Chem. 2002;277:30792—30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, et al. Regulasyon sa JNK signaling pinaagi sa GSTp. EMBO J. 1999;18:1321�1334.
90. Manevich Y, Feinstein SI, Fisher AB. Ang pagpaaktibo sa antioxidant enzyme 1-CYS peroxiredoxin nanginahanglan glutathionylation nga gihusay sa heterodimerization nga adunay pGST. Proc Natl Acad Sci US A. 2004;101:3780â3785.
91. Bunker VW. Free radicals, antioxidants ug pagkatigulang. Med Lab Sci. 1992;49:299–312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. Systemic oxidative stress ug ang relasyon niini sa edad ug sakit. J Am Geriatr Soc. 1996;44:823–827.
93. Puti E, Shannon JS, Patterson RE. Ang relasyon tali sa bitamina ug
paggamit sa calcium supplement ug colon cancer. Cancer Epidemiol Biomarkers Prev. 1997;6:769–774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Mga mekanismo sa nobela sa natural nga antioxidant compound sa biological nga mga sistema: pagkalambigit sa glutathione ug glutathione-related enzymes. J Nutr Biochem. 2005;16:577–586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Mga pagbag-o sa status sa cardiac glutathione human sa ischemia ug reperfusion. Experientia. 1985;41:42–43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Carotenoid radical chemistry ug antioxidant/pro-oxidant nga mga kabtangan. Arch Biochem Biophys. 2004;430:37–48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Ngano nga nagpaabut kita nga ang mga carotenoid mahimong mga antioxidant sa vivo? Libre nga Radic Res. 1997;26:381–398.
98. Niles RM. Mga agianan sa signal sa retinoid chemoprevention ug pagtambal sa kanser. Mutat Res. 2004;555:81–96.
99. Donato LJ, Noy N. Pagpugong sa pagtubo sa mammary carcinoma pinaagi sa retinoic acid: ang proapoptotic nga mga gene mao ang mga target alang sa retinoic acid receptor ug cellular retinoic acid-binding protein II signaling. Cancer Res. 2005;65:8193–8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, et al. Ang Bcl-2 usa ka yawe nga regulator alang sa retinoic acid-induced apoptotic cell death sa neuroblastoma. Oncogene. 2006;25:5046–5055.
101. Dalton TP, Shertzer HG, Puga A. Regulasyon sa ekspresyon sa gene pinaagi sa reaktibo nga oxygen. Ann Rev Pharmacol Toxicol. 1999;39:67–101.
102. Scandalios JG. Genomic nga mga tubag sa oxidative stress. Sa: Meyers RA, ed. Encyclopedia sa Molecular Cell Biology ug Molecular Medicine. Tomo 5. 2nd ed. Weinheim, Alemanya: Wiley-VCH; 2004: 489–512.
103. Ghosh R, Mitchell DL. Epekto sa kadaot sa oxidative DNA sa mga elemento sa tigpasiugda sa pagbugkos sa transcription factor. Mga Nucleic Acids Res. 1999;27:3213–3218.
104. Marietta C, Gulam H, Brooks PJ. Ang usa ka 8, 50-cyclo-20-deoxyadenosine nga samad sa usa ka TATA box nagpugong sa pagbugkos sa TATA binding protein ug kusog nga pagkunhod sa transkripsyon sa vivo. Pag-ayo sa DNA (Amst). 2002;1:967–975.
105. Jackson AL, Chen R, Loeb LA. Induction sa microsatellite instability
pinaagi sa oxidative DNA kadaot. Proc Natl Acad Sci US A. 1998;95:12468�12473.
106. Caldecott KW. Ang mga interaksyon sa protina-protina sa panahon sa mammalian DNA single-strand break repair. Biochem Soc Trans. 2003;31:247–251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidative DNA kadaot: mekanismo, mutation, ug sakit. FASEB J. 2003;17:1195�1214.
108. Jones PL, Wolffe AP. Mga relasyon tali sa organisasyon sa chromatin ug DNA methylation sa pagtino sa ekspresyon sa gene. Semin Cancer Biol. 1999;9:339–347.
109. Girotti AW. Mga mekanismo sa lipid peroxidation. J Free Radic Biol Med. 1985;1:87–95.
110. Siu GM, Draper HH. Metabolismo sa malonaldehyde sa vivo ug in vitro. Mga lipid. 1982;17:349–355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Posible nga pag-apil sa lipid-peroxidation nga produkto 4-hydroxynonenal sa pagporma sa fluorescent chromolipids. Biochem J. 1986;239:405–409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Ang mga pagbag-o nga nagsalig sa edad sa lebel sa lipid peroxide sa mga tipik sa lipoprotein sa serum sa tawo. J Gerontol. 1984;39:269–272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, ug uban pa. 4- Hydroxynonenal, usa ka aldehydic nga produkto sa membrane lipid peroxidation, nagdaot sa glutamate transport ug mitochondrial function sa synaptosomes. Neuroscience. 1997;806:85–96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Pagpaaktibo sa stress signaling pathways pinaagi sa katapusan nga produkto sa lipid peroxidation. Ang 4-hydroxy-2-nonenal usa ka potensyal nga inducer sa paghimo sa intracellular peroxide. J Biol Chem. 1999;274:2234–2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Pagpaaktibo sa EGF receptor pinaagi sa oxidized LDL. FASEB J. 1998;12:665�671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal nagpalambo sa produksiyon sa fibronectin pinaagi sa IMR-90 nga mga fibroblast sa baga sa tawo nga bahin pinaagi sa pagpaaktibo sa epidermal growth factor receptor-linked extracellular signal- regulated kinase p44/42 nga agianan. Toxicol Appl Pharmacol. 2002;184:127–135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Gipagawas ang 8-isoprostane isip in vivo biomarker sa lung oxidative stress sa mga pasyente nga adunay COPD ug himsog nga mga hinabako. Am J Respir Crit Care Med. 2000;162:1175–1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Epithelial permeability, panghubag, ug stress sa oxidant sa mga kahanginan sa mga hinabako. Am J Respir Crit Care Med. 1999;159:473–479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Dugang nga sulod sa thiobarbituric acid-reaktibo nga mga butang ug hydrogen peroxide sa expired gininhawa condensate sa mga pasyente nga adunay lig-on nga chronic obstructive pulmonary nga sakit: walay mahinungdanong epekto sa pagpanigarilyo. Respirar Med. 1999;93:389–396.
120. Kelly FJ, Mudway IS. Ang oksihenasyon sa protina sa interface sa hangin-baga. Amino Acid. 2003;25:375–396.
121. Dean RT, Roberts CR, Jessup W. Fragmentation sa extracellular ug intracellular polypeptides pinaagi sa free radicals. Prog Clin Biol Res. 1985;180:341–350.
122. Keck RG. Ang paggamit sa t-butyl hydroperoxide isip probe alang sa methionine oxidation sa mga protina. Anal Biochem. 1996;236:56–62.
123. Davies KJ. Ang kadaot sa protina ug pagkadaot sa mga radical sa oxygen. I. Kinatibuk-ang mga aspeto. J Biol Chem. 1987;262:9895–9901.
124. Stadtman ER. Metal ion-catalyzed oxidation sa mga protina: biochemical nga mekanismo ug biological nga mga sangputanan. Libre nga Radic Biol Med.
1990;9:315–325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Ang pagpaaktibo sa yawe nga metabolic enzymes pinaagi sa mixed-function oxidation reactions: posible nga implikasyon sa protina turnover ug pagkatigulang. Proc Natl Acad Sci US A. 1983;80:1521�1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oxidation sa methionine residues sa mga protina: biological nga mga sangputanan. Antioxid Redox Signal. 2003;5:577–582.
127. Stadtman ER, Levine RL. Free radical-mediated oxidation sa libre nga amino acids ug amino acid residues sa mga protina. Amino Acid. 2003;25:207–218.
128. Stadtman ER. Ang oksihenasyon sa protina sa pagkatigulang ug mga sakit nga may kalabutan sa edad. Ann NY Acad Sci. 2001;928:22–38.
129. Shacter E. Pag-ihap ug kamahinungdanon sa oksihenasyon sa protina sa biological nga mga sample. Drug Metab Pin. 2000;32:307–326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Oxidative stress ug cell signaling. Curr Med Chem. 2004;11:1163–1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Vascular endothelial growth factor (VEGF) ug ang mga receptor niini. FASEB J. 1999;13:9–22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, et al. Regulasyon sa reactive-oxygen species nga henerasyon sa fibroblasts ni Rac1. Biochem J. 1996;318:379â382.
133. Sun T, Oberley LW. Redox nga regulasyon sa transcriptional activators. Libre nga Radic Biol Med. 1996;21:335–348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redox nga regulasyon sa c-Jun DNA nga nagbugkos pinaagi sa mabalik nga S-glutathiolation. FASEB J. 1999;13:1481�1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. In situ detection sa S-glutathionylated proteins human sa glutaredoxin-1 catalyzed cysteine derivatization. Biochim Biophys Acta. 2006;1760:380–387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulasyon sa glutaredoxin-1
ekspresyon sa usa ka modelo sa mouse sa allergic airway disease. Am J Respir Cell Mol Biol. 2007;36:147–151.
137. Filomeni G, Rotilio G, Ciriolo MR. Cell signaling ug ang glutathione redox system. Biochem Pharmacol. 2002;64:1057–1064.
138. Pande V, Ramos MJ. Molecular recognition sa 15-deoxydelta (12,14) prostaglandin J(2) pinaagi sa nuclear factor-kappa B ug uban pang cellular proteins. Bioorg Med Chem Lett. 2005;15:4057–4063.
139. Perkins ND. Paghiusa sa mga agianan sa pagsenyas sa cell nga adunay function sa NF-kappaB ug IKK. Nat Rev Mol Cell Biol. 2007;8:49–62.
140. Gilmore TD. Pasiuna sa NF-kappaB: mga magdudula, mga agianan, mga panan-aw. Oncogene. 2006;25:6680–6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Lahi nga mga tahas sa thioredoxin sa cytoplasm ug sa nucleus. Usa ka duha ka lakang nga mekanismo sa redox nga regulasyon sa transcription factor NF-kappaB. J Biol Chem. 1999;274:27891�27897.
142. Purok PA. Papel sa komplemento, chemokines ug regulatory cytokines sa acute nga kadaot sa baga. Ann NY Acad Sci. 1996;796:104–112.
143. Akira S, Kishimoto A. NF-IL6 ug NF-kB sa cytokine gene regulation. Adv Immunol. 1997;65:1–46.
144. Meyer M, Schreck R, Baeuerle PA. Ang H2O2 ug antioxidants adunay kaatbang nga mga epekto sa pagpaaktibo sa NF-kappa B ug AP-1 sa mga intact cells: AP-1 isip secondary antioxidant-responsive factor. EMBO J. 1993;12:2005�2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redox regulasyon sa fos ug jun DNA-pagbugkos nga kalihokan sa vitro. Siyensya. 1990;249:1157–1161.
146. Galter D, Mihm S, Droge W. Lahi nga mga epekto sa glutathione disulphide sa nukleyar nga transcription nga mga hinungdan kB ug ang activator protein-1. Eur J Biochem. 1994;221:639–648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. Ang kalihokan sa transkripsyon sa AP-1 gikontrol sa direktang asosasyon tali sa thioredoxin ug Ref-1. Proc Natl Acad Sci US A. 1997;94: 3633�3638.
Ang Find A Practitioner nga himan sa IFM mao ang pinakadako nga referral network sa Functional Medicine, nga gihimo aron matabangan ang mga pasyente nga makit-an ang mga Functional Medicine practitioner bisan asa sa kalibutan. Ang IFM Certified Practitioners gilista una sa mga resulta sa pagpangita, tungod sa ilang halapad nga edukasyon sa Functional Medicine