
Kitang tanan gitudloan isip mga bata nga adunay 5 ka sentido: panan-aw, lami, tingog, baho, ug paghikap. Ang unang upat ka sentido naggamit ug tin-aw, lahi nga mga organo, sama sa mata, taste bud, dalunggan, ug ilong, apan sa unsang paagi nga ang sentido sa lawas motandog gayod? Ang paghikap masinati sa tibuok lawas, sa sulod ug sa gawas. Wala'y usa ka lahi nga organo nga responsable sa paghikap sa paghikap. Hinunoa, adunay gagmay nga mga receptor, o nerve endings, sa palibot sa tibuok lawas nga makamatikod sa paghikap diin kini mahitabo ug magpadala ug mga signal ngadto sa utok uban ang impormasyon bahin sa matang sa paghikap nga nahitabo. Ingon nga ang usa ka taste bud sa dila makamatikod sa lami, ang mga mechanoreceptor maoy mga glandula sulod sa panit ug sa ubang mga organo nga makamatikod sa mga pagbati sa paghikap. Nailhan sila nga mechanoreceptors tungod kay kini gidisenyo aron makamatikod sa mekanikal nga mga sensasyon o mga kalainan sa pressure.
Kaundan
Papel sa Mechanoreceptors
Ang usa ka tawo nakasabut nga sila nakasinati og usa ka pagbati sa higayon nga ang organ nga responsable sa pagdiskubre sa maong piho nga pagbati nagpadala ug mensahe ngadto sa utok, nga mao ang nag-unang organ nga nagproseso ug naghan-ay sa tanang impormasyon. Ang mga mensahe gipadala gikan sa tanang bahin sa lawas ngadto sa utok pinaagi sa mga alambre nga gitawag ug mga neuron. Adunay libu-libo nga gagmay nga mga neuron nga nagsanga sa tanan nga bahin sa lawas sa tawo, ug sa mga tumoy sa kadaghanan niini nga mga neuron adunay mga mechanoreceptor. Aron ipakita kung unsa ang mahitabo kung imong gihikap ang usa ka butang, mogamit kami usa ka pananglitan.
Hunahunaa ang usa ka lamok nga motugpa sa imong bukton. Ang strain niini nga insekto, nga gaan kaayo, makapadasig sa mga mechanoreceptor nianang partikular nga bahin sa bukton. Kadtong mga mechanoreceptor nagpadala ug mensahe subay sa neuron nga ilang konektado. Ang neuron nagkonektar hangtod sa utok, nga nakadawat sa mensahe nga adunay butang nga nagtandog sa imong lawas sa eksaktong lokasyon sa piho nga mechanoreceptor nga nagpadala sa mensahe. Ang utok molihok uban niini nga tambag. Tingali kini mosulti sa mga mata sa pagtan-aw sa rehiyon sa bukton nga nakamatikod sa pirma. Ug kung ang mga mata mosulti sa utok nga adunay lamok sa bukton, ang utok mahimong mosulti sa kamot sa pagdali dayon niini. Ingon niana ang pagtrabaho sa mga mechanoreceptor. Ang katuyoan sa artikulo sa ubos mao ang pagpakita ingon man paghisgot sa detalye sa functional nga organisasyon ug mga determinant sa molekula sa mga mechanoreceptor.
Touch Sense: Functional Organization ug Molecular Determinants sa Mechanosensitive Receptors
abstract
Ang mga cutaneous mechanoreceptors na-localize sa lain-laing mga layer sa panit diin sila makamatikod sa usa ka halapad nga mekanikal nga stimuli, lakip na ang light brush, stretch, vibration ug noxious pressure. Kini nga lainlain nga stimuli gipares sa usa ka lainlain nga han-ay sa mga espesyal nga mechanoreceptor nga motubag sa deformation sa panit sa usa ka piho nga paagi ug ipasa kini nga stimuli sa mas taas nga istruktura sa utok. Ang mga pagtuon sa tibuuk nga mga mechanoreceptor ug genetically tractable sensory nerve endings nagsugod sa pagbutyag sa mga mekanismo sa pagbati sa paghikap. Ang pagtrabaho niini nga natad naghatag sa mga tigdukiduki og mas bug-os nga pagsabot sa organisasyon sa sirkito nga nagpahipi sa panglantaw sa paghikap. Ang mga bag-ong channel sa ion mitumaw ingon nga mga kandidato alang sa mga molekula sa transduction ug mga kabtangan sa mekanikal nga gated nga mga sulog nga nagpauswag sa among pagsabut sa mga mekanismo sa pagpahiangay sa tactile stimuli. Gipasiugda niini nga pagrepaso ang pag-uswag nga nahimo sa pag-ila sa mga functional nga kabtangan sa mga mechanoreceptor sa balhiboon ug glabrous nga panit ug ion nga mga channel nga nakamatikod sa mekanikal nga mga input ug naghulma sa mechanoreceptor adaptation.
keywords: mechanoreceptor, mechanosensitive channel, kasakit, panit, somatosensory system, paghikap
Pasiuna
Ang paghikap mao ang pagkakita sa mekanikal nga stimulus nga nakaapekto sa panit, lakip ang dili makadaot ug makadaot nga mekanikal nga stimuli. Kini usa ka hinungdanon nga pagbati alang sa pagkaluwas ug pag-uswag sa mga mammal ug tawo. Ang pagkontak sa solid nga mga butang ug mga pluwido sa panit naghatag sa gikinahanglan nga impormasyon sa sentral nga sistema sa nerbiyos nga nagtugot sa eksplorasyon ug pag-ila sa palibot ug nagpasiugda sa locomotion o giplano nga paglihok sa kamot. Ang paghikap importante usab alang sa apprenticeship, social contacts ug sexuality. Ang pagbati sa paghikap mao ang labing gamay nga huyang nga pagbati, bisan kung mahimo kini nga gituis (hyperesthesia, hypoesthesia) sa daghang mga kondisyon sa pathological.1-3
Ang mga tubag sa paghikap naglakip sa usa ka tukma kaayo nga coding sa mekanikal nga impormasyon. Ang cutaneous mechanoreceptors na-localize sa lain-laing mga lut-od sa panit diin sila makamatikod sa usa ka halapad nga mekanikal nga stimuli, lakip na ang light brush, stretch, vibration, deflection sa buhok ug noxious pressure. Kini nga lainlain nga stimuli gipares sa usa ka lainlain nga han-ay sa mga espesyal nga mechanoreceptor nga motubag sa deformation sa panit sa usa ka piho nga paagi ug ipasa kini nga stimuli sa mas taas nga istruktura sa utok. Ang mga somatosensory neuron sa panit nahulog sa duha ka grupo: low-threshold mechanoreceptors (LTMRs) nga motubag sa benign pressure ug high-threshold mechanoreceptors (HTMRs) nga motubag sa makadaot nga mekanikal nga stimulation. Ang LTMR ug HTMR cell body nagpuyo sulod sa dorsal root ganglia (DRG) ug cranial sensory ganglia (trigeminal ganglia). Ang mga lanot sa nerbiyos nga may kalabotan sa LTMRs ug HTMRs giklasipikar nga A?-, A?- o C-fibers base sa ilang action potential conduction velocities. Ang mga lanot sa C walay myelinated ug adunay pinakahinay nga katulin sa pagpadagan (~2 m/s), samtang ang A? ug A? ang mga lanot gaan ug bug-at nga myelinated, nagpakita sa intermediate (~ 12 m / s) ug paspas (~ 20 m / s) nga mga tulin sa pagpadagan, matag usa. Ang mga LTMR giklasipikar usab nga hinay, o paspas nga pagpahiangay sa mga tubag (SA- ug RA-LTMR) sumala sa ilang mga rate sa pagpahiangay sa padayon nga mekanikal nga stimulus. Mas gipalahi sila pinaagi sa cutaneous end organs nga ilang gisudlan ug sa ilang gusto nga stimuli.
Ang katakus sa mga mechanoreceptor nga makit-an ang mga mekanikal nga cues nagsalig sa presensya sa mga channel sa mechanotransducer ion nga paspas nga nagbag-o sa mga mekanikal nga pwersa sa mga signal sa kuryente ug nag-depolarise sa natad sa pagdawat. Kining lokal nga depolarization, nga gitawag og receptor potential, makamugna og action potentials nga mokaylap paingon sa central nervous system. Bisan pa, ang mga kabtangan sa mga molekula nga nagpataliwala sa mechanotransduction ug pagpahiangay sa mga mekanikal nga pwersa nagpabilin nga dili klaro.
Sa kini nga pagrepaso, naghatag kami usa ka kinatibuk-ang pagtan-aw sa mga kabtangan sa mammalian mechanoreceptor sa dili makadaot ug makadaot nga paghikap sa balhiboon ug glabrous nga panit. Gikonsiderar usab namo ang bag-o nga kahibalo mahitungod sa mga kabtangan sa mga mekanikal nga gated nga mga sulog sa pagsulay sa pagpatin-aw sa mekanismo sa pagpahiangay sa mechanoreceptor. Sa katapusan, among girepaso ang bag-o nga pag-uswag nga nahimo sa pag-ila sa mga channel sa ion ug mga kaubang protina nga responsable sa pagmugna sa mechano-gated nga mga sulog.
Dili Makadaot nga Paghikap
Mga LTMR nga Nalambigit sa Follicle sa Buhok
Ang mga follicle sa buhok nagrepresentar sa mga mini-organ nga naghimo sa shaft sa buhok nga nakamatikod sa gaan nga paghikap. Ang mga lanot nga may kalabutan sa mga follicle sa buhok motubag sa paglihok sa buhok ug sa direksyon niini pinaagi sa pagpabuto sa mga tren sa mga potensyal nga aksyon sa pagsugod ug pagtangtang sa stimulus. Sila paspas nga nagpahiangay sa mga receptor.
Iring ug koneho. Sa coat ug rabbit coat, ang mga follicle sa buhok mahimong bahinon sa tulo ka klase sa follicle sa buhok, ang Down hair, ang Guard hair ug ang Tylotrichs. Ang Down hairs (underhair, wool, vellus)4 mao ang pinakadaghan, ang pinakamubo ug pinakanindot nga buhok sa coat. Wavy sila, walay kolor ug migawas sa mga grupo nga duha ngadto sa upat ka buhok gikan sa komon nga orifice sa panit. Ang mga balhibo sa Guard (monotrichs, overhears, tophair)4 gamay nga kurbado, pigmented o walay pigmented, ug nag-inusarang migawas gikan sa mga baba sa ilang mga follicle. Ang mga tylotrich mao ang pinakagamay nga gidaghanon, ang pinakataas ug pinakabaga nga mga buhok.5,6 Sila adunay pigmented o walay pigmented, usahay ang duha ug nag-inusarang mogawas gikan sa usa ka follicle nga gilibotan sa usa ka loop sa capillary blood vessels. Ang mga sensory fiber nga suplay sa usa ka follicle sa buhok nahimutang sa ilawom sa sebaceous gland ug gipasangil sa A? o A?-LTMR lanot.7
Sa suod nga pag-apposition sa down shaft sa buhok, sa ilawom sa lebel sa sebaceous gland mao ang singsing sa lanceolate pilo-Ruffini nga mga tumoy. Kini nga mga sensory nerve endings nahimutang sa usa ka spiral course palibot sa shaft sa buhok sulod sa connective tissue nga nagporma sa follicle sa buhok. Sulod sa follicle sa buhok, adunay usab libre nga mga tumoy sa nerbiyos, ang uban niini nagporma mga mechanoreceptor. Kanunay, ang paghikap sa mga corpuscle (tan-awa ang glabrous nga panit) naglibot sa liog nga rehiyon sa tylotrich follicle.
Ang mga kabtangan sa myelinated nerve endings sa panit nga balhiboon sa iring ug koneho gisusi pag-ayo sa panahon sa 1930-1970 (pagsusi sa Hamann, 1995).8 Talagsaon, si Brown ug Iggo, nagtuon sa 772 ka yunit nga adunay myelinated afferent nerve fibers sa saphenous nerves gikan sa iring. ug koneho, nagklasipikar sa mga tubag sa tulo ka matang sa receptor nga katumbas sa mga lihok sa Down hairs (type D receptors), Guard hair (type G receptors) ug Tylotrich hair (type T receptor).9 Ang tanan nga afferent nerve fiber nga mga tubag gihiusa. sa Rapidly Adapting receptor sa tipo I (RA I) pinaagi sa pagsupak sa Pacinian receptor nga ginganlan og RA II. Ang RA I mechanoreceptors nakamatikod sa tulin sa mekanikal nga stimulus ug adunay hait nga utlanan. Wala sila makamatikod sa mga kalainan sa thermal. Burgess ug uban pa. Gihubit usab ang usa ka paspas nga pagpahiangay sa umahan nga receptor nga labing maayo nga motubag sa pag-stroking sa panit o paglihok sa daghang mga buhok, nga gipahinungod sa pagpukaw sa mga katapusan sa pilo-Ruffini. Walay usa sa tubag sa follicle sa buhok ang gipasangil sa C fiber activity.10
Mga ilaga. Sa dorsal balhiboon nga panit sa mga ilaga, tulo ka dagkong matang sa mga follicle sa buhok ang gihulagway: zigzag (mga 72%), awl/auchene (mga 23%) ug guard o tylotrich (mga 5%).11-14 Zigzag ug Awl/ Ang mga follicle sa buhok sa auchenne naghimo sa mas nipis ug mas mugbo nga mga shaft sa buhok ug nalangkit sa usa ka sebaceous gland. Ang guwardiya o tylotrich nga mga buhok mao ang pinakataas sa mga matang sa follicle sa buhok. Sila gihulagway pinaagi sa usa ka dako nga buhok bombilya nga nakig-uban sa duha ka sebaceous glands. Ang guard ug awl/auchene nga mga buhok gihan-ay sa usa ka iterative, regular nga gilay-on nga pattern samtang ang zigzag nga mga buhok dasok nga nagpuyo sa mga panit nga naglibot sa duha ka dagkong mga follicle sa buhok [Fig. 1 (A1, A2 ug A3)].
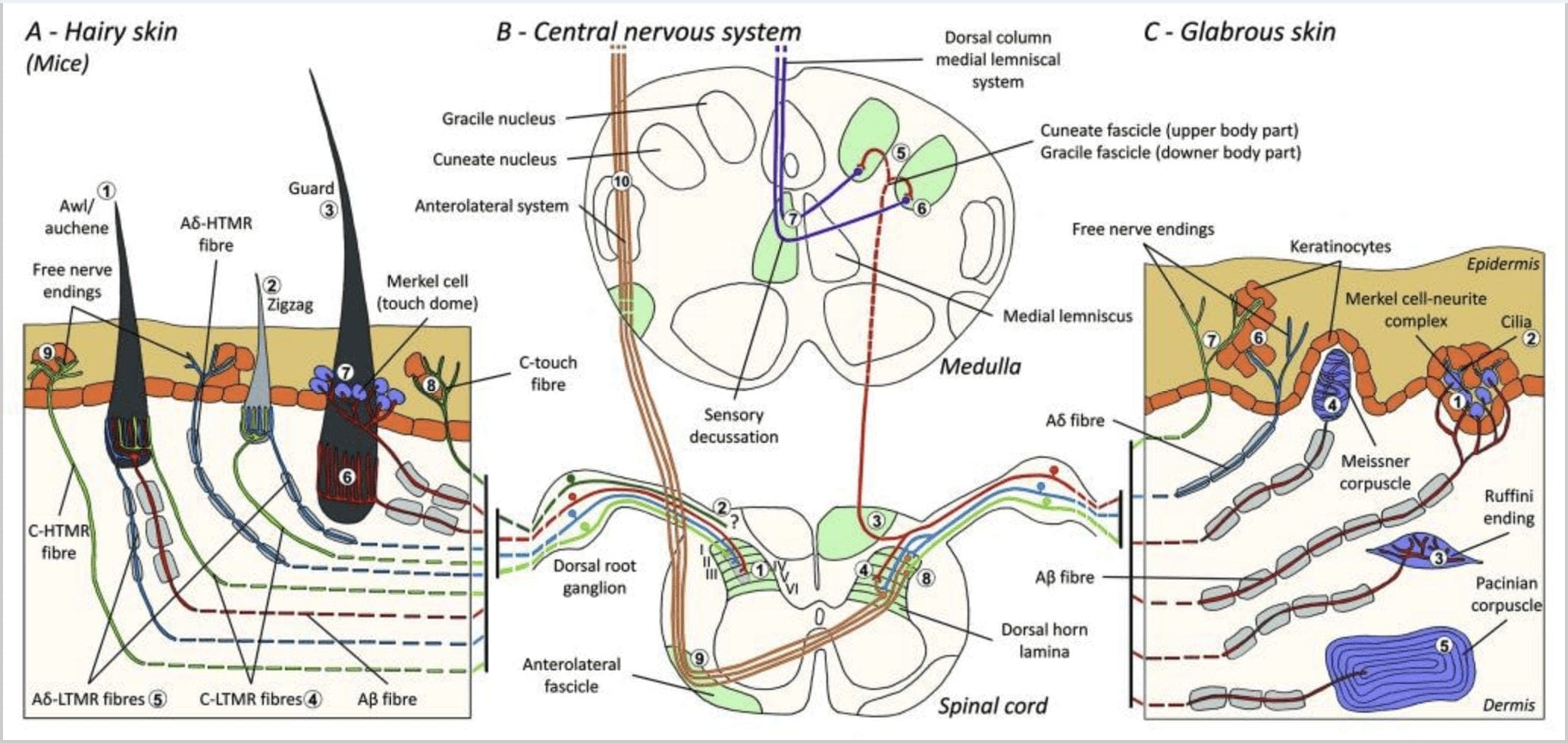
Figure 1. Organisasyon ug mga projection sa cutaneous mechanoreceptors. Sa balhiboon nga panit, ang light brush ug paghikap kasagarang makita sa innervation sa palibot sa mga follicle sa buhok: awl/auchenne (A1), zigzag (A2) ug guard (A3). Ang mga balhibo sa awl/auchene triply innervated sa C-LTMR lanceolate endings (A4), A?-LTMR ug A? paspas nga pagpahiangay-LTMR (A6). Ang zigzag nga mga follicle sa buhok mao ang mas mugbo nga mga shaft sa buhok ug gi-innervate sa C-LTMR (A4) ug A? -LTMR lanceolate endings (A5). Ang pinakataas nga mga follicle sa buhok sa guwardiya gipasulod sa A? paspas nga pagpahiangay-LTMR longitudinal lanceolate endings (A6) ug nalangkit sa A? hinay nga pagpahiangay-LTMR sa touch dome endings (A7). Ang mga sentral nga projection sa tanan niini nga mga lanot natapos sa lahi, apan partially overlapping laminae sa spinal cord dorsal sungay (C-LTMR sa lamina II, A?-LTMR sa lamina III ug A?-LTMR sa lamina IV ug V). Ang mga projection sa LTMR nga nag-innervate sa parehas o kasikbit nga mga follicle sa buhok gilinya aron maporma ang usa ka pig-ot nga kolum sa spinal cord dorsal horn (B1 sa gray). Diha lamang sa balhiboon nga panit, ang usa ka subpopulasyon sa C-fibers free ending makapasulod sa epidermis ug motubag sa nindot nga paghikap (A8). Kini nga mga C-touch fibers dili motubag sa makadaot nga paghikap ug ang ilang agianan sa pagbiyahe wala pa nahibal-an (B2). Sa glabrous nga panit, ang dili makadaot nga paghikap gipataliwad-an sa upat ka matang sa LTMRs. Ang Merkel cell-neurite complex anaa sa basal layer sa epidermis (C1). Kini nga mechanoreceptor naglangkob sa usa ka kahikayan tali sa daghang mga selula sa Merkel ug usa ka gipadako nga terminal sa nerbiyos gikan sa usa ka A? lanot. Ang mga selula sa Merkel nagpakita sa tudlo sama sa mga proseso nga nagkontak sa mga keratinocytes (C2). Ang pagtapos sa Ruffini kay lokal sa mga dermis. Kini usa ka nipis nga pormag tabako nga gisudlan sa mga sensory ending nga konektado sa A? lanot (C3). Ang Meissner corpuscle konektado sa A? nerve ending ug nahimutang sa dermal papillae. Kini nga encapsulated mechanoreceptor naglangkob sa giputos nga suporta nga mga selula nga gihan-ay isip pinahigda nga mga lamellae nga gilibutan sa connective tissue (C4). Ang Pacinian corpuscle mao ang mas lawom nga mechanoreceptor. Usa ka single A? Ang unmyelinated nerve ending natapos sa sentro niining dako nga ovoid corpuscle nga hinimo sa concentric lamellae. Ang mga projection niining A?-LTMR nga mga lanot sa spinal cord gibahin sa duha ka sanga. Ang punoan nga sentral nga sanga (B3) misaka sa spinal cord sa ipsilateral dorsal nga nagporma og cuneate o gracile fascicles (B5) sa lebel sa medulla diin ang mga nag-unang afferent naghimo sa ilang unang synapse (B6). Ang mga sekondaryang neuron naghimo ug sensory decussation (B7) aron maporma ang usa ka tract sa medial lemniscus nga mosaka agi sa brainstem ngadto sa midbrain, ilabina sa thalamus. Ang ikaduhang sanga sa LTMR natapos sa sungay sa dorsal sa lamina II, IV, V ug makabalda sa pagpasa sa kasakit (B4). Ang makahilo nga paghikap makita sa libre nga nerve nga nagtapos sa epidermis sa balhiboon (A9) ug glabrous nga panit (C7). Kini nga mga mechanoreceptor mao ang pagtapos sa A?-HTMR ug C-HTMR sa suod nga kontak sa silingang mga keratinocytes (C6). Ang A?-hTMR matapos sa lamina I ug V; Ang C-HTMR natapos sa lamina I ug II (B8). Sa lebel sa dorsal horn sa spinal cord, ang mga nag-unang afferent nga HTMR naghimo og mga synapses nga adunay mga sekondaryang neuron nga motabok sa midline ug mosaka sa mas taas nga istruktura sa utok sa anterolateral fascicle (B9, B10).
Bag-ohay lang, si Ginty ug mga kauban migamit ug kombinasyon sa molecular-genetic labeling ug somatotopic retrograde tracing approaches aron mahanduraw ang organisasyon sa peripheral ug central axonal endings sa LTMRs sa mga ilaga. gikuha sa tulo ka matang sa follicle sa buhok ug gipasa pinaagi sa mga kalihokan sa talagsaon nga kombinasyon sa A?-, A?- ug C- lanot ngadto sa dorsal horn.
Gipakita nila nga ang genetic labeling sa tyrosine hydroxylase positive (TH+) DRG neurones nagpaila sa usa ka populasyon sa nonpeptidergic, gamay nga diametro nga sensory neurones ug nagtugot sa pagtan-aw sa C-LTMR peripheral endings sa panit. Katingad-an, ang mga sanga sa axoneal sa indibidwal nga mga C-LTMR nakit-an nga nag-arborise ug nagporma sa mga longhitudinal lanceolate nga mga tumoy nga suod nga nakig-uban sa zigzag (80% sa mga katapusan) ug awl / auchene (20% sa mga katapusan), apan dili tylotrich nga mga follicle sa buhok [Fig. 1 (A4)]. Ang mga longhitudinal lanceolate endings dugay na nga gihunahuna nga eksklusibo sa A?-LTMRs ug busa wala damha nga ang mga katapusan sa C-LTMRs maporma nga longhitudinal lanceolate endings.15 Kini nga mga C-LTMR adunay intermediate adaptation kon itandi sa hinay ug paspas nga pagpahiangay. myelinated mechanoreceptors [Fig. 2 (C1)].
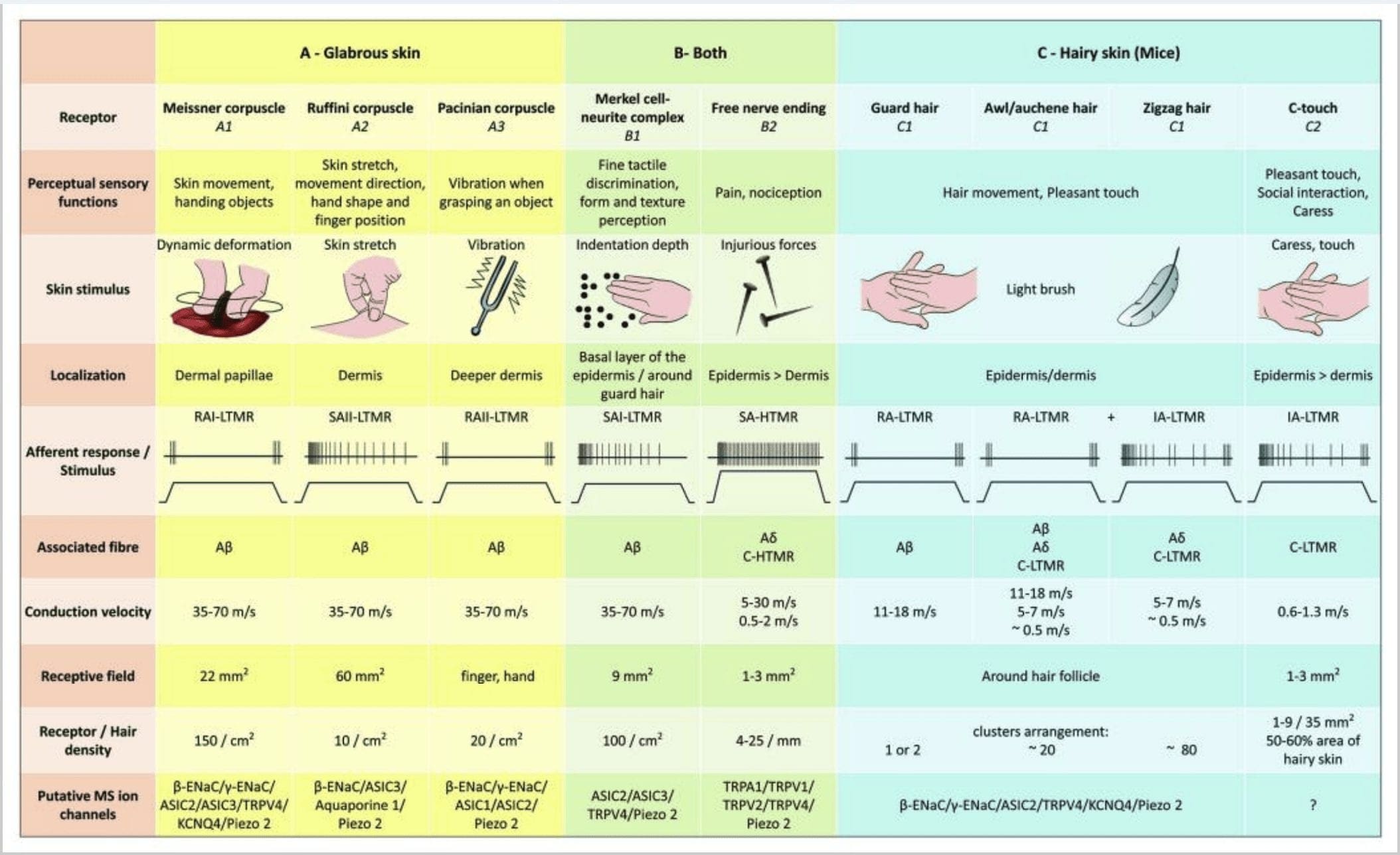
Figure 2. Tactile receptors sa mammals: Ang cutaneous tactile receptors nagkalahi ngadto sa innocuous touch nga gisuportahan sa multiple receptors nga adunay ubos nga mechanical threshold (LTMRs) sa glabrous ug balhiboon nga panit ug noxious touch nga gisuportahan sa high mechanical threshold receptor (HTMRs). Naghimo sila og mga nerve free endings nga nag-una sa epidermis. (A) Glabrous nga panit. A1: Ang Meissner corpuscles nakamatikod sa paglihok sa panit ug sa pagkadait sa butang diha sa kamot. Importante sila sa paghatag og butang ug kaabtik. Ang mga receptor paspas nga mopahiangay sa stimulus, konektado sa A? mga lanot ug gamay sa C lanot ug adunay dako nga natad sa receptor. A2: Ang mga corpuscle sa Ruffini nakamatikod sa pag-inat sa panit ug importante nga makit-an ang posisyon sa tudlo ug butang nga gitunol. Ang receptor hinayhinay nga mopahiangay sa stimulus ug magpadayon nga kalihokan basta ang stimulus gigamit. Ang mga receptor konektado sa A? mga lanot ug adunay dako nga natad sa pagdawat. A3: Ang mga corpuscle sa Pacinian mas lawom sa mga dermis ug nakamatikod sa pagkurog. Ang mga receptor konektado sa A? lanot; sila paspas nga mopahiangay sa stimulus ug adunay pinakadako nga natad sa pagdawat. (B) Tibuok panit. B1: Ang mga Merkel-cell complex anaa sa glabrous nga panit ug sa palibot sa buhok. Sila dasok nga gipahayag sa kamot ug importante alang sa texture perception ug pinakamaayong diskriminasyon tali sa duha ka punto. Sila ang responsable sa katukma sa tudlo. Ang mga receptor konektado sa A? lanot; hinay-hinay silang mopahiangay sa stimulus ug adunay mubo nga natad sa pagdawat. B2: Makadaot nga paghikap sa mga HTMR nga adunay hinay kaayo nga pagpahiangay sa stimulus, ie, aktibo basta ang nociceptive stimulus magamit. Naporma sila sa libre nga nerve ending sa A? ug C-fibers nga nalangkit sa keratinocytes. (C) Balhiboon nga panit. C1: Ang mga follicle sa buhok nalangkit sa lain-laing klase sa buhok. Sa mga ilaga Ang mga buhok sa guwardiya mao ang mas taas ug gamay nga gipahayag, ang awl/auchenne kay medium nga gidak-on ug ang zigzag mao ang pinakagamay ug ang pinakadasok nga gipahayag nga buhok. Konektado sila sa A? mga lanot apan usab sa A? ug C-LTMRs fibers para sa awl/auchenne ug zizag nga buhok. Namatikdan nila ang paglihok sa buhok lakip ang makapahimuot nga paghikap sa panahon sa pag-atiman. Sila paspas nga mopahiangay o uban sa intermediate kinetic sa stimulus. C2: Ang C-touch nerve endings katumbas sa usa ka subtype sa C fibers terminus nga adunay libre nga pagtapos nga gihulagway sa usa ka ubos nga mekanikal nga threshold. Kinahanglan nga mag-encode sila alang sa makapahimuot nga sensasyon nga gipahinabo sa paghaplos. Sila kasarangan nga mopahiangay sa stimulus ug adunay mubo nga natad sa pagdawat. Putative mechanosensitive (MS) ion channels nga gipahayag sa lain-laing mga tactile receptors gipakita sumala sa preliminary data ug summarize present hypothesis ubos sa evaluation.
Ang ikaduha nga mayor nga populasyon nga giila mahitungod sa A?-LTMR endings sa Awl/Auchenne ug zigzag follicles nga itandi sa Down hair follicle nga kaylap nga gitun-an sa iring ug koneho. Gipakita sa Ginty ug mga kolaborator nga ang TrkB gipahayag sa taas nga lebel sa usa ka subset sa medium-diameter nga DRG neurones. Ang mga pagrekord sa intracellular gamit ang ex vivo skin-nerve nga pag-andam sa gimarkahan nga mga lanot nagpakita nga sila nagpakita sa physiological nga mga kabtangan sa mga lanot nga gitun-an kaniadto sa iring ug koneho: talagsaon nga mekanikal nga pagkasensitibo (Von Frey threshold <0.07 mN), paspas nga pagpahiangay sa mga tubag sa suprathreshold stimuli, intermediate conduction velocities (5.8 � 0.9 m/s) ug pig-ot nga wala mausab nga mga spike sa soma.15 Kini nga mga A?-LTMR nagporma ug longhitudinal lanceolate nga mga tumoy nga nalangkit sa halos tanang zigzag ug awl/auchene nga follicle sa buhok sa punoan [Fig. 1 (A5)].
Sa katapusan, gipakita nila nga ang peripheral nga mga katapusan sa paspas nga pagpahiangay sa A? Ang mga LTMR nagporma og longhitudinal lanceolate nga mga tumoy nga may kalabutan sa guard (o tylotrich) ug awl/auchene nga mga follicle sa buhok [Fig. 1 (A6)].15 Dugang pa, ang mga buhok sa Guard nalangkit usab sa usa ka Merkel cell complex nga nagporma og touch dome nga konektado sa A? hinay nga pagpahiangay sa LTMR [Fig. 1 (A7)].
Sa katingbanan, halos tanang zigzag nga mga follicle sa buhok gipasulod sa C-LTMR ug A?-LTMR lanceolate endings; Ang mga buhok sa awl/auchene triply innervated ni A? paspas nga pagpahiangay-LTMR, A?-LTMR ug C-LTMR lanceolate endings; Ang mga follicle sa buhok sa guwardiya gipasulod sa A? paspas nga pagpahiangay-LTMR longhitudinal lanceolate endings ug makig-uban sa A? hinay nga pagpahiangay-LTMR sa paghikap dome endings. Sa ingon, ang matag follicle sa buhok sa ilaga nakadawat talagsaon ug invariant nga mga kombinasyon sa mga pagtapos sa LTMR nga katumbas sa neurophysiologically lahi nga mechanosensory end organs. Sa pagkonsiderar sa iterative nga kahikayan niining tulo ka matang sa buhok, si Ginty ug ang mga kolaborator nagsugyot nga ang balhiboon nga panit naglangkob sa iterative repeat sa peripheral unit nga adunay, (1) usa o duha ka sentral nga nahimutang nga guard hairs, (2) ~20 nga naglibot nga awl/auchenne nga mga buhok ug (3 ) ~80 nga gisal-ot nga zigzag nga mga buhok [Fig. 2 (C1)].
Projection sa spinal cord. Ang sentral nga mga projection sa A? paspas nga pagpahiangay-LTMRs, A?-LTMRs ug C-LTMRs natapos sa lahi, apan partially overlapping laminae (II, III, IV) sa spinal cord dorsal sungay. Dugang pa, ang mga sentral nga terminal sa LTMRs nga nag-innervate sa parehas o kasikbit nga mga follicle sa buhok sa sulod sa usa ka peripheral nga yunit sa LTMR gipahiangay aron maporma ang usa ka pig-ot nga kolum sa LTMR sa spinal cord dorsal horn [Fig. 1 (B1)]. Busa, lagmit nga ang usa ka wedge, o kolum sa somatotopically organized primary sensory afferent endings sa dorsal horn nagrepresentar sa pag-align sa mga sentral nga projection sa A?-, A?- ug C-LTMRs nga nagpasulod sa parehas nga peripheral unit ug nakamatikod sa mekanikal. stimuli nga naglihok sa parehas nga gamay nga grupo sa mga follicle sa buhok. Base sa gidaghanon sa mga guard, awl/auchene ug zigzag nga mga buhok sa punoan ug limbs ug sa mga numero sa matag subtype sa LTMR, gibanabana ni Ginty ug mga kolaborator nga ang sungay sa dorsal sa mouse adunay 2,000-4,000 LTMR columns, nga katumbas sa gibanabana nga gidaghanon sa peripheral Mga yunit sa LTMR.15
Dugang pa, ang mga axone sa LTMR subtypes suod nga nakig-uban sa usag usa, nga adunay entwined projection ug interdigitated lanceolate endings nga nag-innervate sa parehas nga follicle sa buhok. Dugang pa, tungod kay ang tulo ka mga matang sa follicle sa buhok nagpakita sa lain-laing mga porma, gidak-on ug cellular komposisyon, sila lagmit adunay lahi nga deflectional o vibrational tuning kabtangan. Kini nga mga nahibal-an nahiuyon sa klasiko nga mga pagsukod sa neurophysiological sa iring ug koneho nga nagpakita nga ang A? RA-LTMRs ug A?-LTMRs mahimong differentially activate pinaagi sa deflection sa lahi nga buhok follicle matang.16,17
Sa konklusyon, ang paghikap sa balhiboon nga panit mao ang kombinasyon sa: (1) ang relatibong mga numero, talagsaon nga spatial distribution ug lahi nga morphological ug deflectional nga mga kabtangan sa tulo ka matang sa mga follicle sa buhok; (2) ang talagsaon nga mga kombinasyon sa LTMR subtype endings nga nalangkit sa matag usa sa tulo ka matang sa follicle sa buhok; ug (3) lahi nga pagkasensitibo, katulin sa pagpadagan, spike train patterns ug adaptation properties sa upat ka nag-unang klase sa hair-follicle-associated LTMRs nga makapahimo sa balhiboon nga skin mechanosensory system sa pagkuha ug pagpasa ngadto sa CNS sa komplikadong kombinasyon sa mga kalidad nga naghubit sa usa ka paghikap.
Mga Free-Nerve Endings LTMRs
Kasagaran, ang mga C-fibers nga libre nga mga tumoy sa panit mao ang mga HTMR, apan ang usa ka subpopulasyon sa mga C-fibers dili motubag sa makadaot nga paghikap. Kini nga subset sa tactile C-fiber (CT) afferents nagrepresentar sa usa ka lahi nga tipo sa unmyelinated, low-threshold mechanoreceptive nga mga yunit nga anaa sa balhiboon apan dili glabrous nga panit sa mga tawo ug mammal [Fig. 1 (A8)] .18,19 Ang mga CT sa kasagaran nalangkit sa panglantaw sa makapahimuot nga tactile stimulation sa kontak sa lawas.20,21
Ang mga CT afferent motubag sa mga pwersa sa indentation sa range nga 0.3-2.5 mN ug sa ingon sensitibo sa deformation sa panit sama sa kadaghanan sa A? afferents.19 Ang adaptation nga mga kinaiya sa CT afferents mao ang intermediate kon itandi sa hinay ug paspas nga pagpahiangay sa myelinated mechanoreceptors. Ang madawat nga mga natad sa mga CT afferent sa tawo halos lingin o oval ang porma. Ang umahan naglangkob sa usa ngadto sa siyam ka gagmay nga responsive spots nga gipang-apod-apod sa usa ka lugar hangtod sa 35 mm2.22 Ang mouse homolog receptors giorganisar sa usa ka pattern sa discontinuous patches nga naglangkob sa mga 50-60% sa lugar sa balhiboon nga panit [Fig. 2 (C2)].23
Ang ebidensya gikan sa mga pasyente nga kulang sa myelinated tactile afferents nagpakita nga ang pagsenyas sa CT fibers nagpalihok sa insular cortex. Tungod kay kini nga sistema dili maayo sa pag-encode sa diskriminatibo nga mga aspeto sa paghikap, apan haum kaayo sa pag-encode sa hinay, malumo nga paghikap, ang CT fibers sa balhiboon nga panit mahimong kabahin sa usa ka sistema sa pagproseso sa makapahimuot ug sosyal nga may kalabutan nga mga aspeto sa paghikap.24 Ang CT fiber activation mahimo usab adunay papel sa pagpugong sa kasakit ug bag-o lang gisugyot nga ang panghubag o trauma mahimong makausab sa sensasyon nga gipasa sa C-fiber LTMRs gikan sa nindot nga paghikap ngadto sa kasakit.25,26
Unsang agianan ang pagbiyahe sa CT-afferent wala pa mahibal-an [Fig. 1 (B2)], apan ang ubos nga threshold tactile inputs ngadto sa spinothalamic projection cells na-dokumentado, 27 nagpahulam sa pagsalig sa mga taho sa maliputon, contralateral nga mga kakulangan sa touch detection sa mga pasyente sa tawo human sa pagkaguba niini nga mga agianan human sa chordotomy procedures.28
Mga LTMR sa Glabrous nga Panit
Ang Merkel cell-neurite complex ug touch dome. Ang Merkel (1875) mao ang una nga naghatag usa ka histological nga paghulagway sa mga pungpong sa mga selula sa epidermal nga adunay dagkong lobulated nuclei, nga nakigkontak sa gituohan nga afferent nerve fibers. Nagtuo siya nga ilang gipaubos ang pagbati sa paghikap pinaagi sa pagtawag kanila og Tastzellen (tactile cell). Sa mga tawo, ang Merkel cell neurite complex gipadato sa sensitibo nga mga lugar sa panit, kini makita sa basal layer sa epidermis sa mga tudlo, ngabil ug mga kinatawo. Anaa usab sila sa balhiboon nga panit sa ubos nga densidad. Ang Merkel cell neurite complex naglangkob sa usa ka Merkel cell sa duol nga pag-apposition sa usa ka gipadako nga nerve terminal gikan sa usa ka myelinated A? lanot [Fig. 1 (C1)] (pagrepaso sa Halata ug mga kolaborator).29 Sa epidermal nga bahin sa Merkel cell nagpakita sa mga proseso nga sama sa tudlo nga naglugway tali sa silingang mga keratinocytes [Fig. 1 (C2)]. Ang mga selula sa Merkel kay mga selula sa epidermal nga nakuha sa keratinocyte.30,31 Ang termino sa touch dome gipaila aron hinganlan ang dako nga konsentrasyon sa mga Merkel cell complex sa balhiboon nga panit sa forepaw sa iring. Ang usa ka touch dome mahimong adunay hangtod sa 150 ka mga Merkel nga mga selula nga gipasulod sa usa ka A?-fiber ug sa mga tawo gawas sa A?-fibers, A? ug ang C-fibers kanunay usab nga anaa.32-34
Ang pagpukaw sa Merkel cell neurite complex moresulta sa hinay-hinay nga pagpahiangay sa Type I (SA I) nga mga tubag, nga naggikan sa punctuate receptive field nga adunay hait nga mga utlanan. Walay spontaneous discharge. Kini nga mga complex mosanong sa indentation nga giladmon sa panit ug adunay pinakataas nga spatial resolution (0.5 mm) sa cutaneous mechanoreceptors. Nagpadala sila usa ka tukma nga spatial nga imahe sa tactile stimuli ug gisugyot nga mahimong responsable sa pagpihig sa porma ug texture [Fig. 2 (B1)]. Ang mga ilaga nga wala sa Merkel nga mga selula dili makamatikod sa mga textured surface gamit ang ilang mga tiil samtang ilang gibuhat kini gamit ang ilang mga bungot.35
Kung ang Merkel cell, ang sensory neuron o pareho nga mga site sa mechanotransduction usa gihapon ka debate. Sa mga ilaga, ang phototoxic nga pagkaguba sa mga selula sa Merkel nagwagtang sa SA I nga tubag.36 Sa mga ilaga nga adunay genetically suppressed-Merkel cells, ang SA I nga tubag nga natala sa ex vivo nga panit/nerve nga pagpangandam hingpit nga nawala, nga nagpakita nga ang Merkel nga mga selula gikinahanglan alang sa husto nga pag-encode sa Merkel mga tubag sa receptor.37 Bisan pa, ang mekanikal nga pagpadasig sa nahilit nga mga selula sa Merkel sa kultura pinaagi sa presyur nga gimaneho sa motor dili makamugna og mga agianan nga adunay mekanikal nga gate.38,39 Ang mga keratinocytes mahimo’g adunay hinungdanon nga papel sa normal nga paglihok sa Merkel cell neurite complex. Ang Merkel cell nga sama sa tudlo nga mga proseso mahimong molihok uban ang deformation sa panit ug paglihok sa epidermis cell, ug kini mahimong ang unang lakang sa mekanikal nga transduction. Klaro, ang mga kondisyon nga gikinahanglan sa pagtuon sa mechano-sensitivity sa Merkel cells wala pa matukod.
Mga katapusan sa Ruffini. Ang mga tumoy sa Ruffini mga manipis nga pormag tabako nga gisudlan sa mga sensory ending nga konektado sa A? mga tumoy sa nerbiyos. Ang mga tumoy sa Ruffini mao ang gagmay nga connective tissue cylinders nga gihan-ay subay sa dermal collagen strands nga gisuplay sa usa ngadto sa tulo ka myelinated nerve fibers nga 4-6 �m diametro. Hangtod sa tulo ka mga silindro nga lainlain ang oryentasyon sa mga dermis mahimong maghiusa aron mahimong usa ka receptor [Fig. 1 (C3)]. Sa estruktura, ang Ruffini endings susama sa Golgi tendon organs. Kini kaylap nga gipahayag sa mga dermis ug giila nga hinay-hinay nga pagpahiangay sa type II (SA II) nga mga mechanoreceptor sa panit. Batok sa background sa kusog nga kalihokan sa nerbiyos, ang usa ka hinay nga pagpahiangay nga regular nga paggawas makuha pinaagi sa perpendicular low force nga gipadayon ang mekanikal nga pagpukaw o labi ka epektibo pinaagi sa dermal stretch. Ang tubag sa SA II naggikan sa dagkong madawat nga mga uma nga adunay dili klaro nga mga utlanan. Ang mga receptor sa Ruffini nakatampo sa pagsabot sa direksyon sa paglihok sa butang pinaagi sa sumbanan sa pag-inat sa panit [Fig. 2 (A2)].
Sa mga ilaga, ang SA I ug SA II nga mga tubag mahimong mabulag electrophysiologically sa ex-vivo nerve-skin preparation.40 Nandasena ug mga kolaborator nagtaho sa immunolocalization sa aquaporin 1 (AQP1) sa periodontal Ruffini endings sa rat incisors nga nagsugyot nga ang AQP1 nalangkit sa ang pagmentinar sa dental osmotic balance nga gikinahanglan alang sa mechanotransduction.41 Ang periodontal Ruffini endings usab nagpahayag sa putative mechanosensitive ion channel ASIC3.42
Ang Meissner corpuscles. Ang mga corpuscle sa Meissner na-localize sa dermal papillae sa glabrous nga panit, kasagaran sa mga palad sa kamot ug mga lapalapa sa tiil apan usab sa mga ngabil, sa dila, sa nawong, sa mga utong ug sa mga kinatawo. Anatomically, kini naglangkob sa usa ka encapsulated nerve ending, ang kapsula gihimo sa mga flattened supportive cells nga gihan-ay ingong pinahigda nga lamellae nga nasulod sa connective tissue. Adunay usa ka single nerve fiber A? afferent konektado kada corpuscle [Fig. 1 (C4)]. Ang bisan unsang pisikal nga deformation sa corpuscle nagpahinabog usa ka volley sa mga potensyal nga aksyon nga dali nga mohunong, ie, sila paspas nga nagpahiangay sa mga receptor. Sa diha nga ang stimulus gikuha, ang corpuscle mobalik sa iyang porma ug samtang sa pagbuhat sa ingon og laing volley sa aksyon potensyal. Tungod sa ilang taphaw nga lokasyon sa mga dermis, kini nga mga corpuscle mapili nga motubag sa paglihok sa panit, pagtuki sa tactile sa slip ug mga vibrations (20-40 Hz). Sensitibo sila sa dinamikong panit - pananglitan, tali sa panit ug usa ka butang nga gikuptan [Fig. 2 (A1)].
Pacinian corpuscles. Ang mga corpuscle sa Pacinian mao ang mas lawom nga mechanoreceptor sa panit ug mao ang labing sensitibo nga encapsulated cutaneous mechanoreceptor sa paglihok sa panit. Kining dagkong ovoid corpuscles (1 mm ang gitas-on) nga hinimo sa concentric lamellae sa fibrous connective tissue ug fibroblasts nga gilinya sa flat modified Schwann cells gipahayag diha sa lawom nga dermis.43 Sa tunga sa corpuscle, sa usa ka fluid-filled cavity nga gitawag ug inner bulb , nagtapos sa usa ka single A? afferent unmyelinated nerve ending [Fig. 1 (C5)]. Adunay sila usa ka dako nga natad sa pagdawat sa nawong sa panit nga adunay labi ka sensitibo nga sentro. Ang pag-uswag ug pag-obra sa daghang paspas nga pagpahiangay sa mga tipo sa mechanoreceptor nabalda sa c-Maf mutant nga mga ilaga. Sa partikular, ang mga corpuscle sa Pacinian grabe nga na-atrophy.44
Ang Pacinian corpuscles nagpakita nga paspas kaayo nga pagpahaom isip tubag sa indentation sa panit, ang paspas nga pagpahiangay sa II (RA II) nervous discharge nga makahimo sa pagsunod sa taas nga frequency sa vibratory stimuli, ug nagtugot sa panglantaw sa layo nga mga panghitabo pinaagi sa transmitted vibrations.45 Pacinian corpuscle ang mga afferent motubag sa padayon nga indentasyon nga adunay lumalabay nga kalihokan sa pagsugod ug pag-offset sa stimulus. Gitawag usab sila nga mga acceleration detector tungod kay ilang makita ang mga pagbag-o sa kusog sa stimulus ug, kung ang rate sa pagbag-o sa stimulus mausab (sama sa mahitabo sa mga vibrations), ang ilang tubag mahimong proporsyonal sa kini nga pagbag-o. Ang mga corpuscle sa Pacinian nakamatikod sa mga pagbag-o sa gross nga presyur ug labaw sa tanan nga mga vibrations (150�300 Hz), nga ilang makit-an bisan mga sentimetro ang gilay-on [Fig. 2 (A3)].
Ang tonic nga tubag naobserbahan sa decapsulated Pacinian corpuscle.46 Dugang pa, ang intact nga Pacinian corpuscles motubag uban ang padayon nga kalihokan sa panahon sa kanunay nga indentation stimuli, nga walay pag-usab sa mekanikal nga mga threshold o frequency sa pagtubag kung ang GABA-mediated signaling gibabagan tali sa lamellate glia ug usa ka nerve ending.47 Busa, ang mga non-neuronal nga mga sangkap sa Pacinian corpuscle mahimong adunay duha ka papel sa pagsala sa mekanikal nga stimulus ingon man usab sa pag-modulate sa mga kabtangan sa pagtubag sa sensory neurone.
Mga projection sa spinal cord. Ang mga projection sa A?-LTMRs sa spinal cord gibahin sa duha ka sanga. Ang punoan nga sentral nga sanga misaka sa spinal cord sa ipsilateral dorsal columns ngadto sa cervical level [Fig. 1 (B3)]. Ang mga sekondaryang sanga matapos sa sungay sa dorsal sa laminae IV ug makabalda sa pagpasa sa kasakit, pananglitan. Mahimo kini nga makapahuyang sa kasakit isip bahin sa kontrol sa ganghaan [Fig. 1 (B4)].48
Sa lebel sa cervical, ang mga axone sa punoan nga sanga gibulag sa duha ka mga tract: ang midline tract naglangkob sa gracile fascicle nga nagdala sa impormasyon gikan sa ubos nga katunga sa lawas (mga bitiis ug punoan), ug ang gawas nga tract naglangkob sa cuneate fascicle nga nagdala sa impormasyon gikan sa ibabaw nga bahin. sa lawas (mga bukton ug punoan) [Fig. 1 (B5)].
Ang panguna nga tactile afferent naghimo sa ilang una nga synapse nga adunay ikaduhang order nga mga neuron sa medulla diin ang mga lanot gikan sa matag tract synapse sa usa ka nucleus sa parehas nga ngalan: ang gracile fasciculus axones synapse sa gracile nucleus ug ang cuneate axones synapse sa cuneate nucleus [Fig. 1 (B6)]. Ang mga neuron nga nakadawat sa synapse naghatag sa mga sekondaryang afferent ug motabok dayon sa midline aron maporma ang usa ka tract sa contralateral nga bahin sa brainstem—ang medial lemniscus—nga mosaka pinaagi sa brainstem ngadto sa sunod nga relay station sa midbrain, ilabi na, sa thalamus [Fig. . 1 (B7)].
Molekular nga detalye sa LTMRs. Ang mga mekanismo sa molekula nga nagkontrol sa sayo nga pagkalainlain sa mga LTMR bag-o lang nga bahin nga gipatin-aw. Gipakita sa Bourane ug mga kolaborator nga ang mga neuronal nga populasyon nga nagpahayag sa Ret tyrosine kinase receptor (Ret) ug ang co-receptor nga GFR?2 sa E11'13 nga embryonic nga mga ilaga nga DRG gipili nga nagpahayag sa transcription factor nga Mafa.49,50 Kini nga mga awtor nagpakita nga ang Mafa / Ret/GFR?2 neurones nga gitakdang mahimong tulo ka espesipikong mga matang sa LTRMs sa pagkahimugso: ang SA1 neurones nga nag-innervating sa Merkel-cell complex, ang paspas nga pagpahiangay sa mga neuron nga nag-innervating sa Meissner corpuscles ug ang paspas nga pagpahiangay sa mga afferent (RA I) nga nagporma ug lanceolate nga mga tumoy sa palibot sa mga follicle sa buhok. Gitaho usab ni Ginty ug mga kolaborator nga ang mga DRG neuron nga nagpahayag sa sayo nga-Ret paspas nga nagpahiangay sa mga mechanoreceptor gikan sa Meissner corpuscles, Pacinian corpuscles ug lanceolate endings sa palibot sa mga follicle sa buhok.51 Ilang gi-innervate ang mga discrete target zone sa sulod sa gracile ug cuneate nuclei, nga nagpadayag sa usa ka modalory-specisenfic pattern sa mechanocisenfic pattern. neurone axonal projection sulod sa brainstem.
Pagsuhid sa mga mechanoreceptor sa panit sa tawo. Ang teknik sa �microneurography� nga gihulagway ni Hagbarth ug Vallbo niadtong 1968 gipadapat sa pagtuon sa discharge behavior sa single human mechanosensitive endings nga nagsuplay sa muscle, joint ug skin (tan-awa ang review Macefield, 2005).52,53 Ang kadaghanan sa microneurography sa panit sa tawo Gipakita sa mga pagtuon ang pisyolohiya sa mga tactile afferent sa glabrous nga panit sa kamot. Ang mga rekording sa microelectrode gikan sa median ug ulnar nerves sa tawhanong mga sakop nagpadayag sa pagbati sa paghikap nga namugna sa upat ka mga klase sa LTMRs: Ang mga Meissner afferent labi ka sensitibo sa kahayag nga paghapak sa panit, pagtubag sa mga lokal nga pwersa sa paggunting ug nagsugod o dayag nga mga slip sulod sa natad sa pagdawat. Ang mga pacinian afferent hilabihan ka sensitibo sa kusog nga mekanikal nga mga transient. Ang mga afferent kusog nga motubag sa paghuyop sa madawat nga uma. Ang Pacinian corpuscle nga nahimutang sa usa ka digit kasagaran motubag sa pag-tap sa lamesa nga nagsuporta sa bukton. Ang mga afferent sa Merkel sa kinaiyanhon adunay taas nga dinamikong pagkasensitibo sa indentation stimuli nga gigamit sa usa ka discrete nga lugar ug kanunay nga motubag sa usa ka off-discharge sa panahon sa pagpagawas. Bisan kung ang mga afferent sa Ruffini motubag sa mga pwersa nga gigamit nga normal sa panit, usa ka talagsaon nga bahin sa SA II afferent mao ang ilang kapasidad sa pagtubag usab sa lateral nga pag-inat sa panit. Sa katapusan, ang mga yunit sa buhok sa forearm adunay daghang ovoid o dili regular nga pagdawat nga mga uma nga gilangkuban sa daghang mga sensitibo nga lugar nga katumbas sa indibidwal nga mga buhok (matag suplay sa afferent ~ 20 nga buhok).
Mechanical nga pagkasensitibo sa Keratinocytes
Ang bisan unsang mekanikal nga stimulus sa panit kinahanglan nga mapasa pinaagi sa mga keratinocytes nga nagporma sa epidermis. Kini nga mga ubiquitous nga mga selyula mahimo’g maghimo mga function sa pagsenyas dugang sa ilang pagsuporta o pagpanalipod nga mga tahas. Pananglitan, ang mga keratinocytes nagpagawas sa ATP, usa ka importante nga sensory signaling molecule, isip tubag sa mekanikal ug osmotic stimuli.54,55 Ang pagpagawas sa ATP nagduso sa intracellular calcium nga pagtaas pinaagi sa autocrine stimulation sa purinergic receptors.55 Dugang pa, adunay ebidensya nga ang hypotonicity nagpalihok sa Rho -kinase signaling pathway ug ang sunod nga F-actin stress fiber formation nga nagsugyot nga ang mekanikal nga deformation sa mga keratinocytes mahimong mekanikal nga makabalda sa silingang mga selula sama sa Merkel cells alang sa dili makadaot nga paghikap ug C-fiber free endings alang sa makadaot nga paghikap [Fig. 1 (C6)].56,57
Makadaot nga Paghikap
Ang taas nga threshold mechanoreceptors (HTMRs) mao ang epidermal C- ug A? libre nga nerve-endings. Wala sila nakig-uban sa mga espesyal nga istruktura ug naobserbahan sa parehas nga balhibo nga panit [Fig. 1 (A9)] ug glabrous nga panit [Fig. 1(C7)]. Bisan pa, ang termino sa libre nga pagtapos sa nerbiyos kinahanglan nga konsiderahon nga mabinantayon tungod kay ang mga tumoy sa nerbiyos kanunay nga duol sa keratinocyte o Langherans cell o melanocytes. Ang ultrastructural analysis sa nerve endings nagpadayag sa presensya sa rough endoplasmic reticulum, abundant mitochondria ug dense-core vesicle. Ang kasikbit nga mga lamad sa mga selula sa epidermal gipalapot ug susama sa post-synaptic membrane sa mga tisyu sa nerbiyos. Timan-i nga ang interaksyon tali sa nerve endings ug epidermal cells mahimong bidirectional tungod kay ang epidermal cells mahimong magpagawas sa mga mediator sama sa ATP, interleukin (IL6, IL10) ug bradykinin ug sukwahi sa peptidergic nerve endings mahimong magpagawas sa mga peptides sama sa CGRP o substance P nga naglihok sa epidermal cells. Ang mga HTMR naglangkob sa mechano-nociceptors nga gihinam-hinam lamang sa makadaot nga mekanikal nga stimuli ug polymodal nociceptors nga motubag usab sa makadaot nga kainit ug exogenous nga kemikal [Fig. 2 (B2)].58
Ang HTMR afferent fibers natapos sa projection neurones sa dorsal horn sa spinal cord. Ang A?-HTMRs nakigkontak sa ikaduhang order nga mga neuron nga kasagaran sa lamina I ug V, samtang ang C-HTMRs natapos sa lamina II [Fig. 1 (B8)]. Ikaduha nga order nociceptive neurones proyekto sa controlateral nga bahin sa spinal cord ug mosaka sa puti nga butang, nga nagporma sa anterolateral nga sistema. Kini nga mga neuron nagtapos labi na sa thalamus [Fig. 1 (B9 ug B10)].
Mechano-Currents sa Somatosensory Neurones
Ang mga mekanismo sa hinay o paspas nga pagpahiangay sa mga mechanoreceptor wala pa gipatin-aw. Dili klaro kung unsa ang gidak-on sa mechanoreceptor adaptation nga gihatag sa cellular environment sa sensory nerve ending, ang intrinsic nga mga kabtangan sa mechanically-gated channels ug ang mga kabtangan sa axonal voltage-gated ion channels sa sensory neurones (Fig. 2). Bisan pa, ang bag-o nga pag-uswag sa pag-ila sa mekanikal nga gated nga mga sulog nagpakita nga ang lainlaing mga klase sa mga mechanosensitive nga mga kanal anaa sa DRG neurones ug mahimong ipasabut ang pipila ka mga aspeto sa pagpahiangay sa mga mechanoreceptor.
Ang in vitro recording sa mga ilaga nagpakita nga ang soma sa DRG neurons kay intrinsically mechanosensitive ug nagpahayag sa cationic mechano-gated nga mga sulog.59-64 Gadolinium ug ruthenium pula bug-os nga nagbabag sa mechanosensitive nga mga sulog, samtang ang external calcium ug magnesium, sa physiological concentrations, ingon man amiloride ug benzamil, hinungdan sa partial block.60,62,63 FM1-43 naglihok isip usa ka malungtarong blocker, ug ang pag-injection sa FM1-43 ngadto sa pangulahiang tiil sa mga ilaga makapakunhod sa pagkasensitibo sa kasakit sa Randall'Selitto nga pagsulay ug nagdugang sa paw withdrawal threshold nga gisusi. uban sa mga buhok ni von Frey.65
Agig tubag sa padayon nga mekanikal nga pagpadasig, ang mga mechanosensitive nga sulog mikunhod pinaagi sa pagsira. Pinasukad sa mga kanunay nga oras sa pagkadunot karon, upat ka lahi nga lahi sa mga mechanosensitive nga mga sulog ang nahibal-an: paspas nga pagpahiangay sa mga sulog (~3-6 ms), intermediate nga pagpahiangay sa mga sulog (~15-30 ms), hinay nga pagpahiangay sa mga sulog (~200-300 ms). ) ug ultra-hinay-hinay nga pagpahiangay sa mga sulog (~1000 ms).64 Kining tanan nga mga sulog anaa uban ang lain-laing insidente sa mga ilaga DRG neurones nga nagpasulod sa glabrous nga panit sa hindpaw.64
Ang mekanikal nga pagkasensitibo sa mechanosensitive nga mga sulog mahimong matino pinaagi sa paggamit sa usa ka serye sa incremental nga mekanikal nga stimuli, nga nagtugot alang sa medyo detalyado nga stimulus-kasamtang pagtuki.66 Ang stimulus-kasamtangan nga relasyon kay kasagaran sigmoidal, ug ang pinakataas nga amplitude sa kasamtangan gitino pinaagi sa gidaghanon sa mga kanal nga dungan nga bukas.64,67 Makapainteres, ang paspas nga pagpahiangay sa mechanosensitive nga kasamtangan gikataho nga nagpakita sa ubos nga mekanikal nga threshold ug tunga nga pagpaaktibo sa tunga-tunga kon itandi sa ultra-hinay nga pagpahiangay sa mechanosensitive nga kasamtangan.63,65
Ang sensory neurones nga adunay non-nociceptive phenotypes mas gusto nga ipahayag ang paspas nga pagpahiangay sa mechanosensitive nga mga sulog nga adunay ubos nga mekanikal nga threshold.60,61,63,64,68 Sa laing bahin, ang hinay ug ultra-hinay nga pagpahiangay sa mechanosensitive nga mga sulog usahay gitaho sa putative non-nociceptive cells.64,68 Nag-aghat kini nga sugyot nga kini nga mga sulog mahimong makatampo sa lainlaing mga mekanikal nga threshold nga makita sa mga LTMR ug HTMR sa vivo. Bisan tuod kini nga mga in vitro nga mga eksperimento kinahanglan nga himoon uban ang pag-amping, ang suporta alang sa presensya sa soma sa DRG neurones sa ubos ug taas nga threshold mechanotransducers gihatag usab sa radial stretch-based stimulation sa cultured mouse sensory neurones.69 Kini nga paradigm nagpadayag sa duha nag-unang populasyon sa stretch-sensitive neurones, usa nga mosanong sa ubos nga stimulus amplitude ug lain nga pilion nga motubag sa taas nga stimulus amplitude.
Kini nga mga resulta adunay importante, apan espekulatibo, mekanikal nga mga implikasyon: ang mekanikal nga threshold sa sensory neurones tingali adunay gamay nga kalambigitan sa cellular nga organisasyon sa mechanoreceptor apan mahimong anaa sa mga kabtangan sa mechanically-gated ion channels.
Ang mga mekanismo nga nagpailalom sa desensitization sa mechanosensitive cation currents sa ilaga DRG neurones bag-o lang nabungkag.64,67 Kini resulta sa duha ka dungan nga mekanismo nga makaapekto sa channel properties: adaptation ug inactivation. Ang pagpahiangay una nga gitaho sa auditory hair cell nga mga pagtuon. Mahimo kining gihulagway nga operational isip usa ka yano nga paghubad sa activation curve sa transducer channel subay sa mechanical stimulus axis.70-72 Adaptation nagtugot sa sensory receptors sa pagpadayon sa ilang pagkasensitibo sa bag-ong stimuli sa presensya sa usa ka kasamtangan nga stimulus. Bisan pa, ang usa ka dako nga tipik sa mga mechanosensitive nga mga sulog sa DRG neurones dili mahimong ma-reactivate human sa pagkondisyon sa mekanikal nga stimulation, nga nagpakita sa dili aktibo nga pipila ka mga transducer channels.64,67 Busa, ang duha nga inactivation ug adaptation molihok sa tandem aron makontrol ang mechanosensitive nga mga sulog. Kining duha ka mga mekanismo komon sa tanang mechanosensitive nga mga sulog nga giila sa ilaga DRG neurones, nga nagsugyot nga ang mga may kalabutan nga physicochemical nga mga elemento nagtino sa kinetics niini nga mga channel.64
Sa konklusyon, ang pagtino sa mga kabtangan sa endogenous mechanosensitive nga mga sulog sa vitro hinungdanon sa pagpangita sa pag-ila sa mga mekanismo sa transduction sa lebel sa molekula. Ang kabag-ohan nga nakita sa mekanikal nga threshold ug ang pagpahiangay sa mga kinetics sa lainlaing mga mekanikal nga gated nga mga sulog sa DRG neurones nagsugyot nga ang mga intrinsic nga kabtangan sa mga channel sa ion mahimong magpatin-aw, labing menos sa bahin, mekanikal nga threshold ug adaptation kinetics sa mga mechanoreceptor nga gihulagway sa mga dekada 1960. 80 gamit ang ex vivo nga pagpangandam.
Putative Mechanosensitive nga mga Protina
Ang mechanosensitive ion nga mga sulog sa somatosensory neurones maayo nga gihulagway, sa kasukwahi, gamay ra ang nahibal-an bahin sa pagkatawo sa mga molekula nga nagpataliwala sa mechanotransduction sa mga mammal. Ang mga genetic screen sa Drosophila ug C. elegans nakaila sa mga kandidato nga mechanotransduction nga mga molekula, lakip na ang TRP ug degenerin/epithelial Na+ channel (Deg/ENaC) nga mga pamilya.73 Ang bag-o nga mga pagsulay sa pagpatin-aw sa molekula nga basehan sa mechanotransduction sa mga mammal nagpunting sa mga homolog niini nga mga kandidato. . Dugang pa, daghan niini nga mga kandidato anaa sa cutaneous mechanoreceptors ug somatosensory neurones (Fig. 2).
Mga Acid-Sensising Ion Channels
Ang mga ASIC iya sa usa ka proton-gated subgroup sa degenerin�epithelial Na+ channel family.74 Tulo ka membro sa ASIC family (ASIC1, ASIC2 ug ASIC3) ang gipahayag sa mechanoreceptors ug nociceptors. Ang papel sa mga channel sa ASIC gisusi sa mga pagtuon sa pamatasan gamit ang mga ilaga nga adunay gipunting nga pagtangtang sa mga gene sa ASIC channel. Ang pagtangtang sa ASIC1 dili makausab sa function sa cutaneous mechanoreceptors apan nagdugang sa mekanikal nga pagkasensitibo sa mga afferent nga nagpasulod sa gut.75 ASIC2 knockout nga mga ilaga nagpakita sa usa ka pagkunhod sa pagkasensitibo sa paspas nga pagpahiangay sa cutaneous LTMRs.76 Apan, ang misunod nga mga pagtuon nagtaho sa kakulang sa mga epekto sa pag-knockout sa ASIC2 sa pareho nga visceral mechano-nociception ug cutaneous mechanosensation.77 Ang ASIC3 disruption nagpamenos sa mechano sensitivity sa visceral afferents ug nagpamenos sa mga tubag sa cutaneous HTMRs sa noxious stimuli.76
Ang Transient Receptor Channel
ANG TRP superfamily gibahin ngadto sa unom ka subfamilies sa mammals.78 Hapit tanang TRP subfamilies adunay mga miyembro nga nalambigit sa mechanosensation sa lain-laing mga cell system.79 Sa mammalian sensory neurones, bisan pa niana, ang TRP channels labing nailhan tungod sa pag-ila sa thermal nga impormasyon ug pagpataliwala sa neurogenic inflammation, ug duha lang ka TRP channels, TRPV4 ug TRPA1, ang nalambigit sa touch responsiveness. Ang pagsamok sa TRPV4 nga ekspresyon sa mga ilaga adunay kasarangan lamang nga mga epekto sa acute mechanosensory thresholds, apan kusganon nga nakunhuran ang pagkasensitibo sa makadaot nga mekanikal nga stimuli.80,81 TRPV4 usa ka hinungdanon nga determinant sa pagporma sa tubag sa nociceptive neurones sa osmotic stress ug sa mekanikal nga hyperalgesia sa panahon sa panghubag.82,83, 1 TRPA1 daw adunay papel sa mekanikal nga hyperalgesia. Ang mga ilaga nga kulang sa TRPA1 nagpakita sa sobrang pagkasensitibo sa kasakit. Ang TRPA84,85 nakatampo sa transduction sa mekanikal, bugnaw ug kemikal nga stimuli sa nociceptor sensory neurones apan makita nga dili kinahanglanon alang sa transduction sa selula sa buhok.XNUMX
Walay klaro nga ebidensya nga nagpakita nga ang TRP channels ug ASICs channels nga gipahayag sa mammals mechanically gated. Walay usa niini nga mga kanal nga gipahayag nga heterologously nga nag-recapulate sa electrical signature sa mechanosensitive nga mga sulog nga naobserbahan sa ilang lumad nga palibot. Wala niini gisalikway ang posibilidad nga ang mga ASIC ug TRPs nga mga kanal kay mga mechanotransducers, tungod sa kawalay kasiguruhan kung ang usa ka channel sa mechanotransduction mahimong molihok sa gawas sa konteksto sa cellular (tan-awa ang seksyon sa SLP3).
Piezo nga mga protina
Ang mga piezo protiens bag-o lang giila nga mga promising candidates alang sa mechanosensing proteins ni Coste ug collaborators.86,87 Vertebrates adunay duha ka mga miyembro sa Piezo, Piezo 1 ug Piezo 2, kaniadto nailhan nga FAM38A ug FAM38B, matag usa, nga maayo nga gitipigan sa tibuok multi cellular eukaryotes . Ang Piezo 2 abunda sa DRGs, samtang ang Piezo 1 halos dili makita. Ang piezo-induced mechanosensitive nga mga sulog gipugngan nga gipugngan sa gadolinium, ruthenium pula ug GsMTx4 (usa ka hilo gikan sa tarantula Grammostola spatulata).88 Ang ekspresyon sa Piezo 1 o Piezo 2 sa heterologous nga mga sistema nagpatunghag mechanosensitive nga mga sulog, ang kinetics sa inactivation sa Piesterzo2 mao ang faster. kay sa Piezo 1. Susama sa endogenous mechanosensitive nga mga sulog, ang mga sulog nga nagsalig sa Piezo adunay mga potensyal sa pagbag-o sa palibot sa 0 mV ug wala’y gipili nga kation, nga adunay Na +, K +, Ca2 + ug Mg2 + ang tanan nga mituhop sa ilawom nga agianan. Ingon usab, ang mga sulog nga nagsalig sa piezo gikontrol sa potensyal sa lamad, nga adunay marka nga paghinay sa karon nga mga kinetics sa mga potensyal nga depolarized.86
Ang mga protina sa Piezo walay duhaduha nga nag-mechanosensing sa mga protina ug nag-ambit sa daghang mga kabtangan sa paspas nga pagpahiangay sa mga mechanosensitive nga sulog sa sensory neuron. Ang pagtambal sa mga kultura nga DRG neurones nga adunay Piezo 2 nga mubo nga interfering RNA mikunhod ang proporsyon sa mga neuron nga adunay paspas nga pagpahiangay sa kasamtangan ug mikunhod ang porsyento sa mga mechanosensitive neurones.86 Ang mga transmembrane nga domain nahimutang sa tibuok piezo nga mga protina apan walay klaro nga pore-containing motifs o ion channel signatures nga giila. Bisan pa, ang mouse nga Piezo 1 nga protina giputli ug gi-reconstituted ngadto sa asymmetric lipid bilayers ug liposome forms ion channels nga sensitibo sa ruthenium red.87 Usa ka importante nga lakang sa pag-validate sa mechanotransduction pinaagi sa Piezo channels mao ang paggamit sa in vivo approaches aron mahibal-an ang functional importansya sa touch signaling. Gihatag ang kasayuran sa Drosophila diin ang pagtangtang sa usa ka miyembro sa Piezo nagpamenos sa mekanikal nga tubag sa makadaot nga stimuli, nga wala makaapekto sa normal nga paghikap.89 Bisan kung ang ilang istruktura nagpabilin nga determinado, kini nga nobela nga pamilya sa mga mechanosensitive nga protina usa ka maayong hilisgutan alang sa umaabot nga panukiduki, lapas sa utlanan. sa pagbati sa paghikap. Pananglitan, ang bag-o nga pagtuon sa mga pasyente nga adunay anemia (hereditary xerocytosis) nagpakita sa papel sa Piezo 1 sa pagmintinar sa erythrocyte volume homeostasis.90
Sama sa Transmembrane Channel (TMC)
Ang usa ka bag-o nga pagtuon nagpakita nga ang duha ka protina, TMC1 ug TMC2, gikinahanglan alang sa buhok cell mechanotransduction.91 Ang hereditary deafness tungod sa TMC1 gene mutation gitaho sa tawo ug ilaga.92,93 Ang presensya niini nga mga channel wala pa gipakita sa somatosensory system , pero daw maayo ni nga lead para imbestigahan.
Sama sa Stomatin nga Protein 3 (SLP3)
Dugang pa sa mga channel sa transduction, ang pipila ka mga accessory nga protina nga nalambigit sa channel gipakita nga adunay papel sa pagkasensitibo sa paghikap. Ang SLP3 gipahayag sa mammalian DRG neurones. Ang mga pagtuon gamit ang mutant nga mga ilaga nga kulang sa SLP3 nagpakita sa pagbag-o sa mechanosensation ug mechanosentive nga mga sulog.94,95 SLP3 nga tukma nga function nagpabilin nga wala mahibal-an. Kini mahimo nga usa ka linker tali sa mechanosensitive channel ug sa nagpahiping microtubule, ingon sa gisugyot alang sa iyang C. elegans homolog MEC2.96 Bag-ohay lang GR. Ang Lewin lab nagsugyot nga ang usa ka tether gi-synthesize sa DRG sensory neurones ug nagsumpay sa mechanosensitive ion channel ngadto sa extracellular matrix.97 Ang pagkaguba sa link nagwagtang sa RA-mechanosensitive nga kasamtangan nga nagsugyot nga ang pipila ka mga ion channel kay mechanosensitive lamang kung gitali. Ang RA-mechanosensitive nga mga sulog gipugngan usab sa laminin-332, usa ka matrix nga protina nga gihimo sa mga keratinocytes, nga nagpalig-on sa hypothesis sa usa ka modulasyon sa mechanosensitive nga kasamtangan pinaagi sa extracellular proteins.98
K+ Channel Subfamily
Subay sa cationic depolarizing mechanosensitive nga mga sulog, ang presensya sa repolarizing mechanosensitive K+ nga mga sulog gisusi. Ang mga channel sa K + sa mga mechanosensitive nga mga selyula mahimong makalihok sa kasamtangan nga balanse ug makatampo sa pagtino sa mekanikal nga threshold ug sa dagan sa panahon sa pagpahiangay sa mga mechanoreceptor.
Ang mga miyembro sa KCNK nahisakop sa two-pore domain K+ channel (K2P) nga pamilya.99,100 Ang K2P nagpakita sa usa ka talagsaon nga han-ay sa regulasyon pinaagi sa cellular, pisikal ug pharmacological nga mga ahente, lakip ang mga pagbag-o sa pH, kainit, pag-inat ug deformation sa lamad. Kini nga mga K2P aktibo sa pagpahulay sa potensyal sa lamad. Daghang mga subunit sa KCNK ang gipahayag sa somatosensory neurones.101 KCNK2 (TREK-1), KCNK4 (TRAAK) ug TREK-2 nga mga kanal usa sa pipila ka mga agianan diin ang usa ka direkta nga mekanikal nga gating pinaagi sa pag-inat sa lamad gipakita.102,103
Ang mga ilaga nga adunay disrupted nga KCNK2 gene nagpakita sa usa ka gipalambo nga pagkasensitibo sa kainit ug malumo nga mekanikal nga stimuli apan usa ka normal nga withdrawal threshold sa makadaot nga mekanikal nga presyur nga gigamit sa hindpaw gamit ang Randall'Selitto test.104 Ang kulang sa KCNK2 nga mga ilaga nagpakita usab og dugang nga thermal ug mechanical hyperalgesia sa makapahubag. kondisyon. Ang KCNK4 knockout nga mga ilaga sobra ka sensitibo sa malumo nga mekanikal nga pagpadasig, ug kini nga sobrang pagkasensitibo nadugangan pinaagi sa dugang nga dili aktibo sa KCNK2.105 Ang dugang nga mechanosensitivity niining mga knockout nga mga ilaga mahimong magpasabot nga ang pag-inat kasagarang nagpalihok sa depolarizing ug repolarizing mechanosensitive nga mga sulog sa usa ka koordinado nga paagi, susama sa dili balanse nga paagi. depolarizing ug repolarizing boltahe-gated nga mga sulog.
Ang KCNK18 (TRESK) usa ka mayor nga kontribyutor sa background K + conductance nga nag-regulate sa resting membrane nga potensyal sa somatosensory neurones.106 Bisan tuod wala mahibal-an kung ang KCNK18 direkta nga sensitibo sa mekanikal nga stimulation, kini mahimong adunay papel sa pagpataliwala sa mga tubag sa light touch, ingon man masakit nga mekanikal nga pagpukaw. Ang KCNK18 ug sa usa ka gamay nga gidak-on nga KCNK3, gisugyot nga mahimong target sa molekula sa hydroxy-?-sanshool, usa ka compound nga nakit-an sa Schezuan peppercorns nga nagpalihok sa mga receptor sa paghikap ug nagpahinabog tingling sa mga tawo.107,108
Ang boltahe nga nagsalig sa K+ channel KCNQ4 (Kv7.4) hinungdanon alang sa pagtakda sa tulin ug frequency nga gusto sa usa ka subpopulasyon sa paspas nga pagpahiangay sa mga mechanoreceptor sa mga ilaga ug mga tawo. Ang mutation sa KCNQ4 sa sinugdan nalangkit sa usa ka porma sa hereditary deafness. Makapainteres nga ang usa ka bag-o nga pagtuon nag-localize sa KCNQ4 sa peripheral nerve endings sa cutaneous paspas nga pagpahiangay sa follicle sa buhok ug Meissner corpuscle. Tungod niini, ang pagkawala sa function sa KCNQ4 nagdala ngadto sa usa ka pinili nga pagpauswag sa pagkasensitibo sa mechanoreceptor sa ubos nga frequency nga vibration. Ilabi na, ang mga tawo nga adunay late-onset nga pagkawala sa pandungog tungod sa dominanteng mutasyon sa KCNQ4 gene nagpakita sa gipauswag nga performance sa pag-ila sa gamay nga amplitude, ubos nga frequency nga vibration.109

Ang Panabut ni Dr. Alex Jimenez
Ang paghikap giisip nga usa sa labing komplikado nga mga igbalati sa lawas sa tawo, labi na tungod kay wala’y piho nga organo nga nagdumala niini. Hinunoa, ang pagbati sa paghikap mahitabo pinaagi sa sensory receptors, nailhan nga mechanoreceptors, nga makita sa tibuok panit ug motubag sa mekanikal nga pressure o pagtuis. Adunay upat ka nag-unang matang sa mga mechanoreceptor sa glabrous, o walay buhok, nga panit sa mammal: lamellar corpuscles, tactile corpuscles, Merkel nerve endings ug bulbous corpuscles. Ang mga mechanoreceptor naglihok aron tugutan ang pag-ila sa paghikap, aron mamonitor ang posisyon sa mga kaunuran, bukog ug mga lutahan, nga nailhan nga proprioception, ug bisan sa pag-ila sa mga tingog ug paglihok sa lawas. Ang pagsabut sa mga mekanismo sa istruktura ug pag-obra niini nga mga mechanoreceptor usa ka sukaranan nga elemento sa paggamit sa mga pagtambal ug mga terapiya alang sa pagdumala sa kasakit.
Panapos
Ang paghikap usa ka komplikado nga pagbati tungod kay kini nagrepresentar sa lain-laing mga tactile nga mga hiyas, nga mao, vibration, porma, texture, kalipay ug kasakit, uban sa lain-laing mga discriminative performances. Hangtud karon, ang mga sulat tali sa usa ka touch-organ ug ang psychophysical nga pagbati kay correlative ug ang klase-specific molekular marker bag-o lang mitumaw. Ang pag-uswag sa mga pagsulay sa rodent nga katumbas sa pagkalainlain sa pamatasan sa paghikap gikinahanglan karon aron mapadali ang pag-ila sa genomics sa umaabot. Ang paggamit sa mga ilaga nga kulang sa piho nga mga subset sa mga matang sa sensory afferent makapadali pag-ayo sa pag-ila sa mga mechanoreceptor ug mga sensory afferent nga mga lanot nga nalangkit sa usa ka partikular nga paagi sa paghikap. Makapainteres, ang usa ka bag-o nga papel nagbukas sa importante nga pangutana sa genetic nga basehan sa mechanosensory nga mga kinaiya sa tawo ug nagsugyot nga ang usa ka gene mutation mahimong negatibo nga makaimpluwensya sa pagkasensitibo sa paghikap.110 Kini nagpasiugda nga ang pathophysiology sa depisit sa paghikap sa tawo sa usa ka dako nga bahin wala mahibal-an ug siguradong sigurado. pag-uswag pinaagi sa pag-ila sa tukma sa subset sa sensory neurones nga nalambigit sa usa ka touch modality o usa ka touch deficit.
Agig balos, ang pag-uswag nahimo aron mahibal-an ang biophysical nga mga kabtangan sa mechano-gated nga mga sulog.64 Ang pag-uswag sa bag-ong mga teknik sa bag-ohay nga mga tuig, nga nagtugot sa pag-monitor sa mga pagbag-o sa tension sa lamad, samtang ang pagrekord sa mechano-gated nga kasamtangan, napamatud-an nga hinungdanon nga pamaagi sa eksperimento aron ihulagway. mechanosensitive nga mga sulog nga adunay paspas, intermediate ug hinay nga pagpahiangay (gisusi sa Delmas ug mga kolaborator).66,111 Ang umaabot mao ang pagtino sa papel sa kasamtangan nga mga kabtangan sa mga mekanismo sa pagpahiangay sa functionally diverse mechanoreceptors ug ang kontribusyon sa mechanosensitive K+ nga mga sulog ngadto sa excitability sa Mga LTMR ug HTMR.
Ang molekular nga kinaiya sa mechano-gated nga mga sulog sa mga mammal usa usab ka umaabot nga maayong hilisgutan sa panukiduki. Ang umaabot nga panukiduki mag-uswag sa duha ka mga panan-aw, una aron mahibal-an ang papel sa molekula nga aksesorya nga nagdugtong sa mga kanal sa cytoskeleton ug kinahanglan nga itugyan o i-regulate ang mechanosensitivity sa mga channel sa ion nga sama sa TRP ug ASIC / EnaC nga mga pamilya. Ikaduha, sa pag-imbestigar sa dako ug promising nga dapit sa kontribusyon sa Piezo channels pinaagi sa pagtubag sa importante nga mga pangutana, kalabot sa permeation ug gating mekanismo, ang subset sa sensory neurones ug paghikap modalities nga naglambigit sa Piezo ug ang papel sa Piezo sa non neuronal mga selula nga nakig-uban sa. mechanosensation.
Ang pagbati sa paghikap, kon itandi nianang sa panan-aw, lami, tingog ug baho, nga naggamit ug espisipikong mga organo sa pagproseso niini nga mga sensasyon, mahimong mahitabo sa tibuok lawas pinaagi sa gagmay nga mga receptor nga nailhang mechanoreceptors. Ang lain-laing mga matang sa mechanoreceptors makita sa lain-laing mga lut-od sa panit, diin sila makamatikod sa usa ka halapad nga han-ay sa mekanikal nga stimulation. Ang artikulo sa ibabaw naghulagway sa piho nga mga highlight nga nagpakita sa pag-uswag sa istruktura ug functional nga mga mekanismo sa mechanoreceptors nga may kalabutan sa pagbati sa paghikap. Ang impormasyon nga gi-refer gikan sa National Center for Biotechnology Information (NCBI). Ang sakup sa among kasayuran limitado sa chiropractic ingon man sa mga kadaot ug kondisyon sa taludtod. Aron hisgutan ang hilisgutan, palihug ayaw pagpangutana kang Dr. Jimenez o kontaka kami sa�915-850-0900�.
Gi-curate ni Dr. Alex Jimenez

Dugang nga mga Hilisgutan: Sakit sa Bukobuko
Sakit sa likod mao ang usa sa labing kaylap nga hinungdan sa pagkabaldado ug wala’y trabaho nga mga adlaw sa trabaho sa tibuuk kalibutan. Sa tinuud, ang sakit sa bukobuko gipasangil nga ikaduha nga kasagarang hinungdan sa mga pagbisita sa opisina sa doktor, nga mas daghan kaysa mga impeksyon sa taas nga respiratoryo. Gibana-bana nga 80 porsyento sa populasyon ang makasinati og usa ka matang sa sakit sa likod labing menos kausa sa tibuok nilang kinabuhi. Ang dugokan usa ka komplikado nga istruktura nga gilangkoban sa mga bukog, lutahan, ligaments ug kaunuran, ug uban pang humok nga mga tisyu. Tungod niini, mga kadaot ug/o nagkagrabe nga mga kondisyon, sama sa herniated discs, mahimong mosangpot sa mga sintomas sa sakit sa likod. Ang mga samad sa sports o mga aksidente sa awto sa kasagaran mao ang labing kanunay nga hinungdan sa sakit sa bukobuko, bisan pa, usahay ang pinakasimple nga mga paglihok mahimong adunay sakit nga mga sangputanan. Maayo na lang, ang alternatibong mga opsyon sa pagtambal, sama sa pag-atiman sa chiropractic, makatabang sa pagpagaan sa sakit sa likod pinaagi sa paggamit sa mga pag-adjust sa taludtod ug mga pagmaniobra sa manwal, nga sa katapusan makapauswag sa paghupay sa kasakit.

DUGANG IMPORTANTE NGA TOPIC: Low Back Pain Management
DUGANG MGA TOPIS: DUGANG DUGANG: �Taga nga Pain & Treatments
Blangko
mga pakisayran
Close Accordion
Propesyonal nga Sakop sa Pagpraktis *
Ang kasayuran dinhi sa "Structural ug Functional Mechanisms sa Mechanoreceptors" wala gituyo nga pulihan ang usa-sa-usa nga relasyon sa usa ka kwalipikado nga propesyonal sa pag-atiman sa kahimsog o lisensyado nga doktor ug dili tambag medikal. Giawhag ka namon nga maghimo mga desisyon sa pag-atiman sa kahimsog base sa imong panukiduki ug pakigtambayayong sa usa ka kwalipikado nga propesyonal sa pag-atiman sa kahimsog.
Impormasyon sa Blog ug Kasangkaran nga mga Panaghisgot
Ang among sakup sa kasayuran limitado sa Chiropractic, musculoskeletal, pisikal nga mga tambal, kaayohan, nga nag-amot sa etiological mga kasamok sa viscerosomatic sulod sa mga klinikal nga presentasyon, kaubang somatovisceral reflex clinical dynamics, subluxation complex, sensitibo nga mga isyu sa panglawas, ug/o functional nga mga artikulo sa medisina, mga hilisgutan, ug mga diskusyon.
Among gihatag ug present klinikal nga kolaborasyon uban sa mga espesyalista gikan sa lainlaing mga disiplina. Ang matag espesyalista gidumala sa ilang propesyonal nga sakup sa praktis ug ilang hurisdiksyon sa lisensya. Gigamit namon ang mga protocol sa kahimsog ug kahimsog aron matambal ug suportahan ang pag-atiman sa mga kadaot o sakit sa musculoskeletal system.
Ang among mga video, mga post, mga hilisgutan, mga hilisgutan, ug mga insight naglangkob sa mga klinikal nga butang, mga isyu, ug mga hilisgutan nga may kalabutan ug direkta o dili direkta nga nagsuporta sa among klinikal nga sakup sa praktis.*
Ang among opisina makatarunganon nga misulay sa paghatag suporta nga mga citation ug nahibal-an ang may kalabutan nga pagtuon sa panukiduki o mga pagtuon nga nagsuporta sa among mga post. Naghatag kami mga kopya sa pagsuporta sa mga pagtuon sa panukiduki nga magamit sa mga board sa pagdumala ug publiko kung gihangyo.
Nasabtan namon nga gisakup namon ang mga butang nga nanginahanglan dugang nga pagpatin-aw kung giunsa kini makatabang sa usa ka piho nga plano sa pag-atiman o protokol sa pagtambal; busa, aron sa dugang nga paghisgot sa hilisgutan sa taas, palihug ayaw pangutana Dr. Alex Jimenez, DC, O kontaka kami sa 915-850-0900.
Ania kami aron matabangan ka ug ang imong pamilya.
Panalangin
Dr. Alex Jimenez D.C., MSACP, RN*, CCST, Ang IFMCP*, CIFM*, ATN*
email: coach@elpasofunctionalmedicine.com
Lisensyado isip Doktor sa Chiropractic (DC) sa Texas & New Mexico*
Lisensya sa Texas DC # TX5807, New Mexico DC Lisensya # NM-DC2182
Lisensyado isip Rehistradong Nars (RN*) in Florida
Lisensya sa Florida nga RN Lisensya # RN9617241 (Kontrol No. 3558029)
Compact nga kahimtang: Multi-State License: Gitugotan sa Pagpraktis sa Mga Estado sa 40*
Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Akong Digital Business Card





